
�}27.1�@�w���O���r�������̐Ғœ����̎�Ԃɂ�����A�~�m�_�z��̈Ⴂ
���͎��ۂ̃f�[�^�ŁA�E�͒������Ɋ�Â��Đ��肵�������ł�
�@���㐶���Ȋw�́A��`�q���琶�Ԍn�܂ł̊e�K�w�ɂ����镨���I��ՁA���Ȃ킿DNA�ƃ^��
�p�N���̕����E���w�I�����Ɛ������ۂƂ̊W�𖾂炩�ɂ��邱�Ƃ�
����āA�����Ɋւ��鑽���̒m���������炵�܂����B�Ƃ�킯�A����ɂ���ē���ꂽ���l�Ȑ����̕��ՓI�`���́A�����Ȋw�ɑ傫�ȃp���_�C���]���������炵�܂�
���B����������ŁA���l���i���j�Ɛi���i���j���j�Ƃ��������_���A�����𗝉������ł����ɏd�v�����ĔF�������錋�ʂƂ��Ȃ�܂����B�{�u�`�̎c��̕�����
�́A����DNA�ƃ^���p�N���Ƃ������ׂĂ̐����̑̂��\�����镁�ՓI�ȕ��q����ՂɁA�������琶�܂�Ă��鑽�l���i���j�Ɛi���i���j���j�̖��ɂ��ĉ��
���Ă������Ǝv���܂��B
�@�݂Ȃ���͐i���Ƃ����Ƃǂ̂悤�Ȃ��Ƃɕ����ׂ�ł��傤���B�_�[�E�B���̐i���_�A�P���Ȑ������畡�G�Ȑ����ցA���邢�͉����Ȑ������獂���Ȑ����� �Ƃ������Ԃ̗���Ȃǂ��܂��܂ł��傤�B������w��������1970�N�㏉�߂���܂ŁA�w��I�ɂ͐i���͂��܂�d�v������Ă��Ȃ������悤�Ɏv���܂��B�������� �i���͒N���������Ƃ�����܂���B�����͂܂��A�����ς牻�����Ƃɂ����c�_�����S�ŁA�����畜�����ꂽ�������݂��ɁA���邢�͌������鐶���Ƃǂ������Ă� �āA�ǂ����Ⴄ�� �Ƃ��������Ȃ��ϓI�Ȏ����������ɂ��Ċw��̌n���\�z����Ă��܂����B�������A�ߋ��ɑ��݂������������ׂĉ��Ƃ��Ďc���Ă���킯�ł͂���܂��A���@ ����Ă���킯�ł�����܂���A�����҂����R�ɑz����c��܂��A�����n���ł��邽�߁A�i���w�͋q�ϐ��Ɍ�����w��ƈʒu�Â����A�����w�҂����͂����� �����ӎU�L�����������A����o���Ȃ��A����ȏɂ������悤�Ɏv���܂��B
�@�Ƃ��낪�A1970�N��ɂ͂���ƁADNA����舵���Z�p���}���ɐi�����A�l���͈�ς��܂����B�Ɖu�ł̗�����i����̔�����1970�N�㔼�ł����A�������� �w�i�Ő���������ꂽ���̂ł����B���͂��̎����ADNA�Ƃ������q���������Ƃ���A�����w�҂≻�w�҂������Ȋw�̕���ɎQ������悤�ɂȂ�ADNA�̉�͂��瓾�� �� �����ʂ����ƂɁA����܂łƂ͈قȂ�V���Ȏ��_����i���𑨂��悤�Ƃ��錤���҂���������Ă��܂����B���̌��ʁA����܂ŕ`���Ă����i���̃C���[�W�͑傫���ς��܂� ���B�����č��ł́A�����Ȋw�̂����Ȃ镪��ł��낤�ƁA�i����O���ɂ����Ȃ��c�_�͂��蓾�Ȃ��Ƃ����ɂ܂łȂ��Ă��܂��B�@���݂̂ɗ����Ă����i���w���ADNA�̒m���邱�Ƃłǂ̂悤�ȏ��邱�Ƃ��ł��A����ɂ���Ăǂ̂悤�ɐi���𑨂���悤�ɂȂ����̂��A�����
�b�����ꂩ�炵�����Ǝv���Ă��܂��B
�@DNA�͐e����q�ւƈ�`����`���镪�q�ł��BDNA��4��ނ̉���AA�AT�AC�AG�̔z��A����z��ɂ���Ă��̏����L�q���Ă��܂��B�����āA���� �͒n����̂��ׂĂ̐������ɋ��ʂł��B���̂��߁A����Q�̐�������r����Ƃ��A�����ڂ̗ގ��_�⑊��_�ł͂Ȃ��A���̉���z����r���邱�Ƃ��ł��܂��B �ɒ[�� ���Ƃ������A�q�g�ƃV�N���������r����A�ƌ����Ă����f���܂����ADNA���x�����������r���邱�Ƃ��\�ƂȂ�̂ł��B
�@��������܂��傤�B���L�̂P�`�V�́A7��ނ̚M���ނɂ��āA����DNA�̉���z��̈ꕔ������ׂĕ\�L�������̂ł��B�ł��邾����������c�ɕ��Ԃ悤�ɂ��� ����A�q�g�̉���z�����Ƃ��āA��v���Ȃ������Ԃŕ\�����Ă��܂��i���Ȃ݂ɁA�����M���ނ�DNA��1�Z�b�g�͂��悻30���������Ȃ�܂��BDNA�� �P�Z�b�g�̓Q�m���Ƃ��܂��B���������āA����̓Q�m���̂ق�̈ꕔ��Ꭶ���������ł��j�B�����z����A���C�������g����Ƃ����܂��B1 AAGCTTCATAGGAGCAACAATACTAATAATCGCAC�@�}�E�X 2 AAGCTACATAGGAGCAACCGCCCTTATGATTGCCC�@�E�V 3 AAGCTTTACAGGTGCAACCGTCCTCATAATCGCCC�@�e�i�K�U�� 4 AAGCTTCACCGGCGCAACCACCCTCATGATTGCCC�@�I�����E�[�^�� 5 AAGCTTCACCGGCGCAGTTGTTCTTATAATTGCCC�@�S���� 6 AAGCTTCACCGGCGCAATTATCCTCATAATCGCCC�@�`���p���W�[ 7 AAGCTTCACCGGCGCAGTCATTCTCATAATCGCCC�@�q�g
�@�ǂ��ł��傤���B����Ȃɂ��݂��Ɏ��Ă���Ǝv�����A����Ȃɂ��Ⴄ�Ǝv�����͎�ςł����A�Ⴂ���������قnj����ڃq�g�Ɏ��Ă��āA�傫���قnj����ڂ̈� �� ���傫���Ȃ�A���ʂ͂����̊��o�����̂܂ܔ��f���Ă���悤�Ɏv���܂��B�������A�������Ⴄ�Ƃ��A��������v���Ă��邩���ʓI�Ɏ������Ƃ��ł��܂��B���̈� ���ɈӖ�������̂Ȃ�A��̈Ⴂ���A��ϓI�ɂł͂Ȃ��A���l�������ċq�ϓI�ɕ\�������ł��B
�@�Ƃ���ŁADNA�ɂ̓^���p�N���̃A�~�m�_�z��̏������Ă��܂����B���������āA�^���p�N���̃A�~�m�_�z����r���邱�Ƃł����l�ȉ�͂��\�� ���B�������A�P�̃A�~�m�_�ɑ��ĕ����̉���z��i�R�h���j���Ή����Ă��܂����̂ŁA���m�ɂ́A����z����r���邱�ƂƃA�~�m�_�z����r���邱�ƂƂ͈� �Ȃ�̂ł����A���ꂩ��̋c�_�́A���̈Ⴂ�ɍ��E�����قnj����Ȃ��̂ł͂Ȃ��A�ߎ��I��͂ł��̂ŁADNA�ƃ^���p�N���̂ǂ���̔z��Ŕ�r���邩�ɑ傫�� �Ⴂ�͐����܂���B
�@�����ō��x�́A�^���p�N�����r������������܂��傤�B
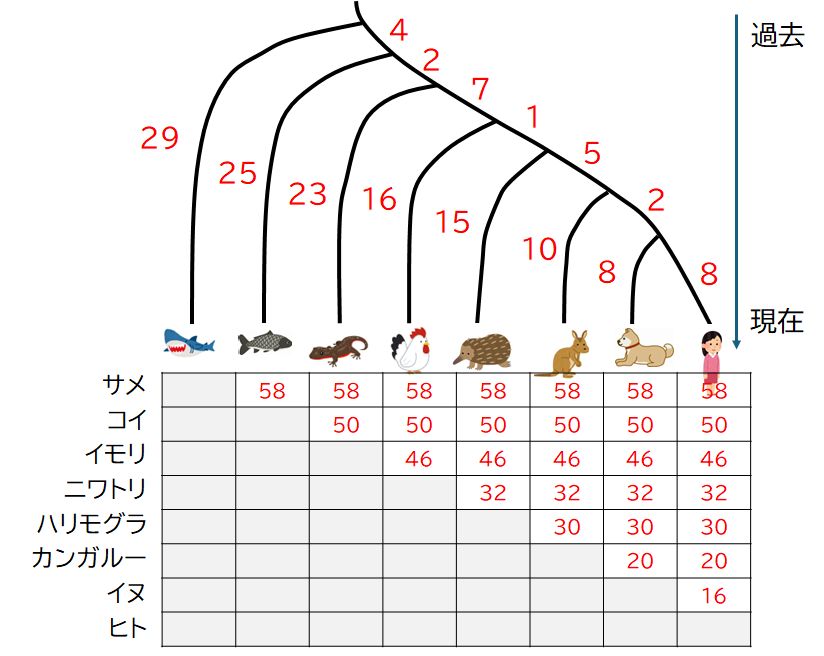
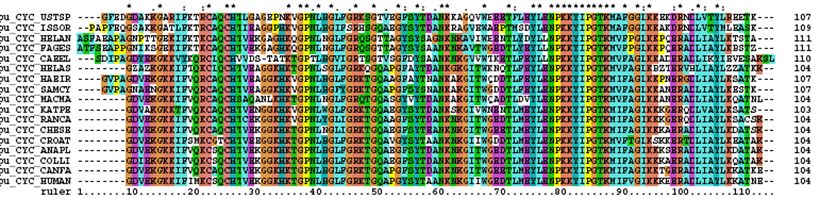
�@�}27.1�́A�����̐Ԍ����ɂ����āA�x���疖���ւƎ_�f���^�ԓ���������w���O���r���̃����Ƃ���^���p�N���̃A�~�m�_�z������낢��ȐҒœ����̎� �ԂŔ� �r�������̂ł��B�Ƃ肠�������̐}�̍����̐��������ɒ��ڂ��Ă������������̂ł����A������2��ԂŃA�~�m�_���قȂ��Ă�ӏ��̊����i%�j�������Ă��܂��B�@�����ƌ���ƁA��ŚM���ނ��r�����悤�ɁA�i���I�ɋ߂���Ԃ̐������������i���Ȃ킿�A���Ă��āj�A�����قǑ傫���Ȃ�i����Ă� ��j�X���ɂ���̂ł����A���ڂ��ׂ��̓T���ƃR�C�ł��B�������ނɑ�����̂ɁA���̈Ⴂ���傫�����܂��H�����ŁA����͒P�Ȃ��O�I�Ȏ��ۂƑ�����ׂ����ƂȂ̂��A�� ��Ƃ��� �炩�̍����I�Ȑ������\�Ȃ̂������ƂȂ�܂��B
�@�T���Ƃ���ȊO�̐Ғœ����Ƃ̈Ⴂ�����Ă݂܂��傤�B�R�C���܂߁A�T��58%�̈Ⴂ������܂��B����ɁA�R�C�ƁA�T������������ȊO�̐Ғœ����Ƃ̈Ⴂ��
�� �Ă݂�ƁA���Ȃ킿�A�R�C���番���Ғœ����ƃR�C�̈Ⴂ�Ƃ������ƂɂȂ�̂ł����A�T��50%�A�C������
�番���Ғœ����ƃC�����Ƃ̈Ⴂ�͊T��46%�ƂȂ��Ă��܂��i�E�̐}�̐Ԏ��̐����ɒ��ڂ��Ă��������I�j�B�ǂ����A�^���p�N���̃A�~�m�_�z��̈Ⴂ�́A
�����̌����ڂȂǂ��画�f�����킪�߂��Ƃ������Ƃ��ł͂Ȃ��A�i���n������ŕ��Ă���̎��Ԃɔ�Ⴕ�Ă���悤�Ɏv���܂��B�������A�킪�߂��Ƃ���
���Ƃ́A���̕��Ă���̎��Ԃ���ʂɂ�
�Z���̂ŁA�킪�߂��ƃA�~�m�_�z���Ă���Ƃ������ƂɂȂ�܂����A�T���ƃR�C�Ƃ����悤�ɁA�������ނł��A�����̎��Ԃ��������̂�����A����������Ԃ̈�
�����������̂́A����܂ł̕��ފw��̎�̋߉����ł͂Ȃ��A����̎��Ԃɒ��ڂ���K�v�����肻���ł��B
�@�������A�T���ƃq�g�̈Ⴂ�̑傫���ƃT���ƃR�C�̂��ꂪ�قړ������Ƃ́A���Ȃ킿�A�^���p�N���̃A�~�m�_�z��i���邢��DNA�̉���z��j�̈Ⴂ�́A��̋�
�����ł͂Ȃ��A�i���n������̕���̎��Ԃɔ�Ⴕ�Ă���Ƃ͉����Ӗ����Ă���̂ł��傤���B���̖₢�������A�i���̑�������傫���ς����d�v�ȃ|�C���g��
��Ȃ̂ł��B
�쐶�^ �@|C T G|G T T|G T C|T A C|C C T|... �@ Leu Val Val Tyr Pro ���`�u���@ |C T G|G T T|G T A|T A C|C C T|... �@ Leu Val Val Tyr Pro �T�C�����g�ψ� �@ ����u�����N�����Ă��@�Ή�����A�~�m�_�ɕω�������܂��� �`�u�� �@|C T G|G T T|G A C|T A C|C C T|... �@ Leu Val Asp Tyr Pro �~�X�Z���X�ψ� ����u�����N�����āA�Ή�����A�~�m�_���ς��܂��� �@ �@�@ |C T G|G T T|G T C|T A A|C C T|... Leu Val Val �I�~ �i���Z���X�ψ� ����u�����N�����āA�I�~�R�h��������܂��� ���� |C T G|G T T|G T T|A C C|C T A|... Leu Val Val Thr Leu �t���[���V�t�g ����������A�z���ɂP�V�t�g�������� ���̌�̉���z�����ƕς���Ă��܂��܂��� �}�� |C T G|G T T|G T A|C T A|C C C|T.. �@ �@ Leu Val Val Leu Pro�@ �t���[���V�t�g ����}������A�z�E�ɂP�V�t�g�������� ���̌�̉���z�����ƕς���Ă��܂��܂���
�@���`�u���̏ꍇ�A�^���p�N�����x���ł̓A�~�m�_�z��ɕω����N����܂���̂ŁA��`�q�Ƃ��Ă͕ς��Ȃ��ƌ����܂��B�������A�`�u���A�����A�}���ł� �^���p�N���̃A�~�m�_�z��ɕω����N����܂��B���̏ꍇ�̉e���͂ǂ��ł��傤���B���낢��ȏꍇ���l�����܂��B�@�L�Q�E�v���I�A�A�@�\�͓������� ���K���I�A�B���K���I�Ȃ܂������V�K�̋@�\�����A�����ćC�قƂ�lje���Ȃ����l�����܂��B�����̉e�����l���Ă�����ő�Ȏ��_�̈�́A���ܐi�� �ɂ��čl���Ă���̂ŁA����DNA�̕ψق��N������1�̂ւ̉e�������ł͂Ȃ��A���ꂪ������ē`�����Ă����ߒ��ŁA���̕ψق��p������q���̌̐� �������邩�ۂ� (������W�c�ւ̌Œ�Ƃ����܂�) ����ɂ��Ȃ���Ȃ�Ȃ��Ƃ������Ƃł��B
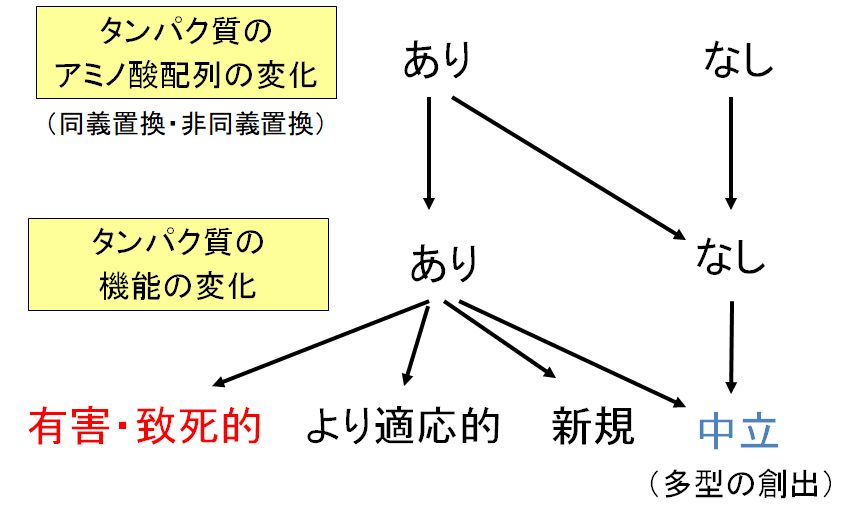

�@�����l����ƁA�@�̗L�Q�E�v���̕ψق��������q���͐i���̉ߒ��œ�������A���ɏ����h�ƂȂ�܂��B�A�ƇB�͐����̗L��������W�c���̑����h�ƂȂ�A��
��
�����m���������ł��傤�B�������A�����I�ɋN����ψقɂ���Ă��K���I�ƂȂ�Ƃ����̂͂����߂����ɂ͋N���肻��������܂���B�C�́A�u�i���ɑ��Ē�
���v�ƕ\������܂����A�ǂ����������Ȃ��̂ŁA�����h�ɂȂ邩�A�����h�ɂȂ邩�͂܂������̊m���ɂ���Č��܂�܂��B�������A�P�ɐ��N�m����������ɂ����
��A�@��������������܂��A����������ւƈ����p�����_���l������ƁA�C�̒����I�ȕψق����|�I�ɑ����ƍl�����܂��B
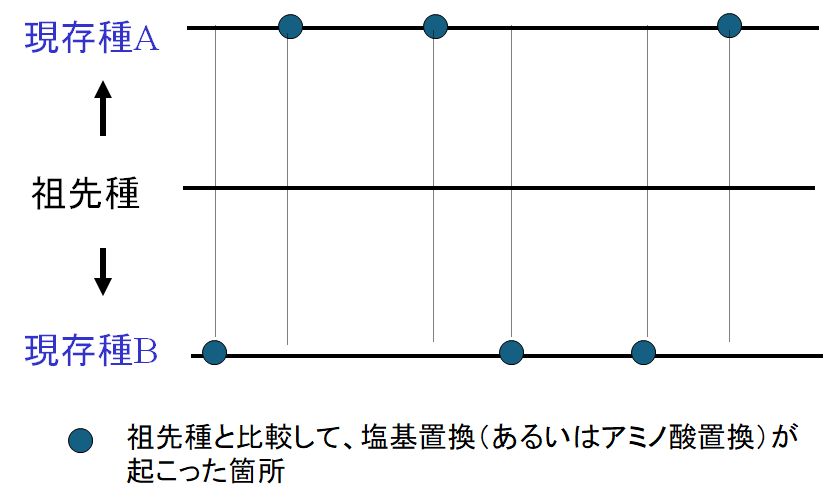
�@�����I�ȕψق̃C���[�W�Ƃ��āA��̑���̈�`�q���l���Ă݂܂��傤�B�炪�e�Ɏ��Ă���Ƃ����̂́A���̈�`�q�������p���ł��邩��ł��B��̑���Ɋւ��
��`�q�͉��S�Ƃ���Ǝv���܂����A��̑��삪�獷���ʂł���̂́A�����̈�`�q�A���Ȃ킿DNA�̉���z��ɏ�ŏq�ׂ��悤�ȈႢ�����邩��ł��B����ł�
�����̎���̐l�X�̊�����Ă��������B�ǂ̊炪�i���I�ɗL���ł��傤���H���ɐ������ɉe������悤�ȈႢ�͂Ȃ������ł��B���ꂪ�i���ɒ����ȕψقł��B���Ȃ�
���A�����ȕψق͑��l���ނ����ŁA�K���I�Ȑi���ɂ͉���v�����Ȃ����̂ł��B���������āA�������`�I���Ƃ�����̂̑����͒����I�ł���A������
���ψق͍ł��p�ɂɂ݂�����̂Ȃ̂ł��B
�@��������DNA�̒����I�ψق̐i���ɂ�����e�����ڍׂɒ��ׁA���̏d�v�������߂Ďw�E�����͓̂��{�̈�`�w�������i�O���s�j�̋����ؑ������ł����B 1967�N�A�u���q���x���iDNA�A�^���p�N�����x���j�̕ω��͑啔�������R�����ɑ��ėL���ł��s���ł��Ȃ������ȕω��ł���A�����I���ۂł���ˑR�ψق� ��`�I�������i���̎�����Ȃ��v�Ƃ��āA����q�i���̒������Ɩ��Â��܂����B�����ň�`�I�����Ƃ́A������ւ̈�`�q�̈��p�����m���I���ۂł��邱�Ƃ��w ���Ă��܂��B
�@�������A�����A���{�ł͐i���w�̑�䏊�A�����юi�̎Љ�w�I�Ȑi���_���S���ł��������ƁA���m�ł́A���������_�[�E�B����ے肵�Ă�����̂Ƃ��Ď� ��ꂽ���Ƃ������āA�Ȃ��Ȃ�������܂���ł����B�������A�ŏ��ɏq�ׂ��悤�ɁA70�N��A80�N���DNA�������g�߂Ȃ��̂ƂȂ�A��̓f�[�^������ ����ɂ�A�����̌����҂̔F�߂���̂ƂȂ�A���݂ł́A�i���w�̒��Ŋm�ł��闝�_�Ƃ��čL���F�߂��Ă��܂��B
�@����������Ȃ��������R�̈���A�������͂��ׂĂ̓ˑR�ψق������ɑ��Ē����ł���Ǝ咣���Ă���ƌ�����ꂽ���Ƃł����B�������A��ŏq�ׂ��悤 �ɁA�L�Q�E�v���ȓˑR�ψق̏o���Ƃ��̓������������ɂƂ��ďd�v�ȑO��ƂȂ��Ă��邱�ƁA�_�[�E�C�����̎��R�I���ɂƂ��ėL���ȓˑR�ψقɂ��Ă��A���̑��� ��ے肷��킯�ł͂Ȃ����Ƃ����������悤�ɂȂ�܂����B�����A�L���ȓˑR�ψق͒����ȓˑR�ψقɔ�ׂ����H�ɂ����N����Ȃ��̂ŁA���̐߂ň������q���v�� ���ĉ�͂���Ƃ��A�ψق̂��ׂĂ������ȕψقł���Ƃ���ߎ���p���邽�߁A�_�[�E�B���̐i���_��ے肵�Ă���悤�ɑ�����ꂽ�̂ł����A�����Ă����ł͂Ȃ� ���Ƃ�ؑ������͍ĎO�������Ă��܂��B
�@�_�[�E�B���̐i���_�ł́A�����ɗL���Ȉ�`�q���W�c�ɍL�܂邱�Ƃ���������܂��B�œK�҂̐����iThe survival of the fittest�j�ł��B����ɑ��āA�ؑ������́A�ǂ����������Ȃ���`�q���W�c�ɍL�܂�̂͊m���̖��ł���A�L�܂����Ƃ���A ����͉^���悩�������炾�Ƃ��āA���� �����ł��^�̂悢�̂��������c�� The survival of the luckiest �ƕ\�����܂����B���̂Q�̗v�������G�ɗ��ݍ����Ȃ���i��ł����̂��i���Ȃ̂ł��B
 �@�@
�@�@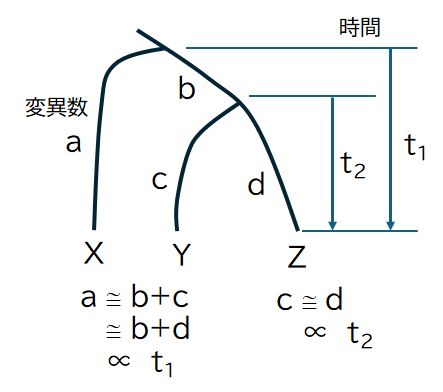
�}27.3�@���ʂ̑c��킩�番���A��A�ɂ���B�ɂ������p�x�ŕψق��N�������Ƃ���ƁA
������̔z����r�����Ƃ��̈Ⴂ�̐��̔����������ɂ��ꂼ��̎�ŋN�������ψق̐��ɑ������A
���̐��͕����̎��Ԃɔ�Ⴗ��Ƃ݂Ȃ����Ƃ��ł��܂��B
�@���āA�}27.1�ɖ߂��Ă݂܂��傤�B�A�~�m�_�̈Ⴂ�̐��̎����l�ɑ��āA�����̕ψِ������̊m���ŋN����Ƃ��āA�����l�ƍ����悤��
���������l���E�ɐԂŎ����܂����B�n������̐Ԃ������́A�n�����̐��̒��������ɔ�Ⴕ�A�����ǂ̂悤�Ȑ�����ƂȂ������Ɉˑ����Ȃ��Ƃ��Đ��肵�Ă����
���B
�����āA�\�ł́A����2�̎���l�����Ƃ��A������q���n������̐��̒����i�ԐF�̐����̘a�j��Ԏ��Ŏ����Ă��܂��B�����l�Ƃ����悻��v���Ă��邱�Ƃ��m�F
�ł���Ǝv���܂��B
�@�������̓I�ȗ�������܂��傤�B�V�g�N���[��C�Ƃ���100�قǂ̃A�~�m�_����Ȃ�^���p�N��������܂��B�قƂ�ǂ̐��������^���p�N���ł��邽
�߁A���܂��܂Ȑ�����ԂŃA�~�m�_�z����r���邱�Ƃ��ł��܂��B�}27.4�ɂ��낢��Ȑ�����̃A�~�m�_�z����A���C�������g�������̂������Ă��܂��B�A�~
�m�_�� 1�����\�L�Ŏ����Ă��܂��B�ڍׂ͓ǂ߂Ȃ��Ƃ��\���܂���B���܂��܂Ȑ����킪�܂܂�Ă��邱�Ƃɒ��ڂ��Ă��������B
CYC_USTSP Ustilago sphaerogena
�i�^�ۗށj�ACYC_ISSOR Issatchenkia orientalis �i�y��j�A
CYC_HELAN Helianthus annuus �i�Ђ܂��j�A CYC_FAGES Fagopyrum esculentum �i���j�A
CYC_CAEEL Caenorhabditis elegans�i�����j�A CYC_HELAS Helix aspersa �i�J�^�c�����j�A
CYC_HAEIR Haematobia irritans �i�n�G�j�A
CYC_SAMCY Samia cynthia �i�J�C�R��j�A
CYC_MACMA Macrobrachium malcolmsonii �i�N���}�G�r�j�ACYC_KATPE Katsuwonus
pelamis �i�J�c�I�j�A
CYC_RANCA Rana catesbeiana �i�E�V�K�G���j�ACYC_CHESE Chelydra serpentina �i�J���j
CYC_CROAT Crotalus atrox �i�K���K���w�r�j�ACYC_ANAPL Anas platyrhynchos �i�A�q���j�A
CYC_COLLI Columba livia �i�n
�g�j�A
CYC_CANFA Canis familiaris �i�C�k�j�A
CYC_HUMAN Homo sapiens �i�q�g�j
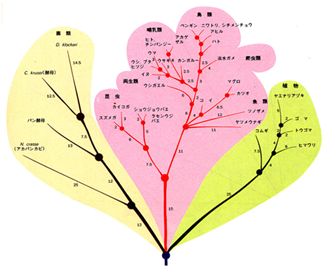
�@�}27.5�E�́A���̕\�����Ƃɍ��ꂽ�n�����ł��B2�̎���q���n������̐��̒������A�~�m�_�z��̈Ⴂ�̐��ɂł��邾����v����悤�ɕ`���ꂽ����
�ł��B��
�����A���w�̕������������悤�ɂ�������Ɖ������܂�킯�ł͂Ȃ��A�\���̂�����͂��������邽�߁A���ꂾ���Ōn�����������ɕ`����Ƃ����킯�ł͂Ȃ��A
���Ȃǂ̒����œ���ꂽ�m���Ƃ��A������̕��މߒ��œ���ꂽ�m���Ȃǂ��Q�l�ɂ��Ē������A�ŏI�I�ɍ\�z����K�v������܂��B
�@������d�v�ȓ_�́A�ψق̐�������̎��Ԃɔ�Ⴗ��킯�ł����A���̔��萔���킩��Ȃ����Ƃł��B�������A���ɏq�ׂ�悤�ɁA���̔��萔�̓^���p
�N�����ƂɈقȂ邽�߁A���ՓI�ȗʂƂ͂Ȃ蓾�Ȃ����Ƃ����ӂ��K�v�ł��B���̂��߁A�]���̉��Ȃǂ̔��@�����Ȃǂ̃f�[�^�����Ƃɐ��肳�ꂽ�n�����Ɣ�r����
����A���ꂼ��̃^���p�N�����Ƃɒ�߂�K�v������܂��B
�@�������Ă��܂��܂ȃ^���p�N���ɂ��ĉ�͂�i�߂����ʁA�����悻���̕��q���v�Ƃ�����͎�@���@�\���邱�ƁA���萔�̓^���p�N�����ƂɈقȂ邱�ƂȂǂ�
�������Ă��܂����B�@�\�I�ɏd�v�łȂ����́i�܂��̓^���p�N���̃A�~�m�_�z���ŏd�v�łȂ������j�قǁA�d�v�Ȃ��́i���邢�̓��p�N���̃A�~�m�_�z���ŏd�v
�ȕ����j���i���̉ߒ��ŃA�~�m�_�i����ёΉ�����DNA�̉���j�̕ψق��p�ɂɋN����i�i�����x���傫���j���Ƃ����炩�ɂȂ�܂����B�d�v�ȋ@�\�����^��
�p�N���قǁA�ψق����̓��ŏd��ȋ@�\��Q�������N�����m���������A��������Ă��܂��A�ψق���e������Ȃ����߂ƍl�����܂��B�܂��A�^���p�N���̃A�~�m
�_�z��̒��ł��A�d�v�ȗ̈�ł̓A�~�m�_�̕ψق͋@�\�s�S���N�����m���������̂ɑ��āA�A�~�m�_�̎�ނɑ��Ă��Ȃ莩�R�x�̍����̈�ł́A�p�ɂɕψق���
�����A���e����Ă��邱�Ƃ��킩���Ă��܂��B
�@����܂ʼn��ɂ̂ݗ����Ă����i���w�́A�������鐶����DNA�̉���z���^���p�N���̃A�~�m�_�z����r���邱�ƂŁA�i����_���邱�Ƃ��\�ƂȂ�܂� ���B��������A�n���̂��ׂĂ̐�����DNA��^���p�N���̔�r�ɂ���ē�����c��ȏ����Ȃ������ł���_���A���ꂪ�i���w�ɋ��߂�����̂ƂȂ��� �̂ł��B
���Ă�H�@���ĂȂ��H�@�`�@�����ڂ̗ގ��x�Ƃc�m�`�̗ގ��x
�V�[���J���X�@�`�@���������ƌĂ�鐶���̃Q�m��