前回の講義の最後に、T細胞受容体と[ペプチド+MHC] 複合体の結合についてお話ししました。それによれば、ヘルパーT細胞あるいはキラーT細胞表面に露出しているT細胞受容体というタンパク質は、その結合部位がT細胞ごとに 異なり、その結合部位の形は1000億種類以上あると推定されています。抗体と同様、遺伝子のランダムな再構成により、これだけの多様性が生み出されていま す。 そして、この膨大なレパートリーによって、樹状細胞が提示する病原体由来の [抗原ペプチド+MHC] 複合体がどのようなものであれ、それにピタッと結合するT細胞受容体をもつT細胞がほぼ確実に存在することになります。樹状細胞が、自己細胞の死骸を食べて、それに 由来する [自己ペプチド+MHC] を提示しても、それにピタッと結合するT細胞受容体をもつT細胞はすでに排除されているため、そうしたT細胞が存在する可能性はほとんどありません。したがって、T細胞受 容体が[ペプチド+MHC] 複合体と結合すれば、そのペプチドは抗原ペプチドである、という論理でした。
今回は、その[自己ペプチド+MHC]複合体と結合するT細胞受容体をもつT細胞排除の経緯と、これまでほとんどその詳細について触れてこなかったMHC というタンパク質についてお話ししようと思います。
誕生したばかりのT細胞、未熟T細胞は自己細胞と非自己細胞を判別する能力を胸腺でチェックされ、合格したものだけが成熟T細胞となり、全身へとその活動 範囲を広げていくことができます。一方、不合格となったT細胞は、アポトーシスへと誘導され、排除されます。
そのチェックが行われる胸腺は、上縦隔に位置するリンパ器官です(図25.1)。生後間もない時 期に体重比では最大となりますが、臓器重量として最大になるのは思春期で、思春期を過ぎると早くも脂肪が入り込んで退縮がはじまります。
胸腺においては、胸腺上皮細胞が、さまざまな [自己ペプチド+MHC]複合体を未熟T細胞に提示し、T細胞受容体の自己ペプチドに対する感受性をテストします(図25.2)。
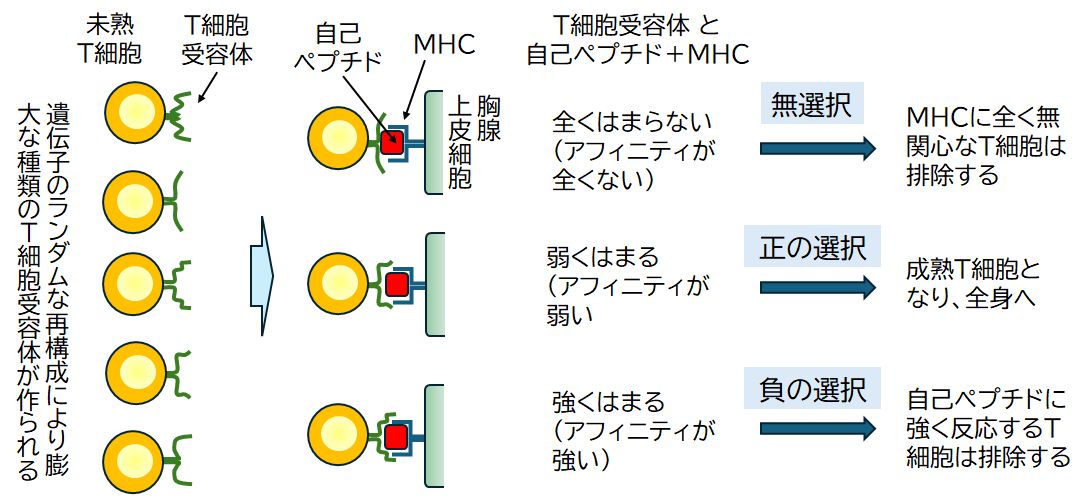
① まず、この複合体に全く結合せず、反応しないT細胞受容体をもつT細胞は死滅させられます。何の役にも立たないT細胞が誤った活動を引き起こすことで何らかの障害を引き起 こすことを避けるためです。② 次に、この複合体と強く結合できるT細胞受容体をもつT細胞も死滅させられます。自己ペプチドを提示する正常細胞を攻撃し、組織障害を起こしてしまうからです。これを負の 選択といいます。③ 結局、この複合体に弱く結合するT細胞受容体をもつT細胞だけが生き残ります。これを正の選択といいます。この選択によって自己のMHCを認識し、かつ自己ペプチドに反応 しないT細胞受容体をもつT細胞だけが生き残ります。
T細胞受容体は、遺伝子の再構成によってランダムに作られるわけですが、こうした選択で大半(98%)のT細胞は死滅させられ、生き残るT細胞は残りの 2% 程度となります。正の選択で残ったT細胞は、T細胞受容体とMHCとの結合を補強する接着剤的な役割を持つタンパク質CD4あるいはCD8を細胞表面に発現させ、 これが合格証の付与となります。CD4が発現したものがヘルパーT細胞、CD8が発現したものがキラーT細胞(細胞障害性T細胞)となります。そして、合格証 を 得たT細胞は胸腺を去り、全身へと送り出されます。
こうして送り出されたT細胞は、[自己ペプチド+MHC]複合体に強く結合することはありませんから、もし[ペプチド+MHC]複合体に強く結合すること が起これば、その複合体に含まれるペプチドは自己のものではなく、非自己、すなわち抗原ペプチドであるということの証明になるわけです。
B細胞受容体(抗体)についてはどうでしょうか。すでに前々回の講義でもお話ししましたが、もう一度確認しておきましょう。B細胞は骨髄で分化成熟します が、その過程で出会うタンパク質はすべて自己由来のものである可能性が高く、それらに反応するB細胞受容体をもつB細胞を除去するという仕組みがあります。ま た、L鎖遺伝子の再構成が再び起こって、自己抗原反応性を回避するメカニズムがあることも見出されています。その他には、脾臓やリンパ節において自己反応性の B細胞を除去する、あるいは機能的に不活性化する仕組みもあります。さらに、前回お話ししたように、B細胞が最終的に活性化されるためには、ヘルパーT細胞に よ るチェックが必要で、自己と非自己の判断は厳密に行われています。
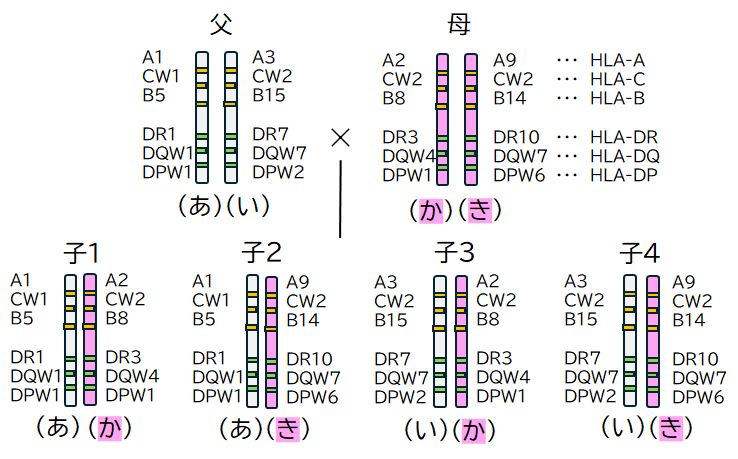
T細胞がT細胞受容体を使ってチェックを行う相手方、MHCについても見ていきましょう。MHCの主要な遺伝子座(DNA上の遺伝子のある場所)は6つあ り、A、B、C、DP、DQ、DRとよばれています。A、B、CがクラスIのMHCの遺伝子座、DP、DQ、DRがクラスIIのMHCの遺伝子座です。実は、 こ の6つの遺伝子座の遺伝子は同じヒトという種に属していても非常に多様性(多型)があることがわかっています。たとえば、Aが30種類ほど、Bが60種類ほ ど、Cが10 種類ほどあることが知られています。両親からそれぞれこれら遺伝子を受け継ぎますから、クラスIの遺伝子座が6、クラスIIの遺伝子座が6あるわけで、それぞれの遺伝子座 にある遺伝子の多様性により、その 組み合わせの場合の数は膨大なものになります。
これらの遺伝子座は、DNA上近くにあるため、ほとんどの場合まとまって親から子へと伝わります。図25.3に例を示しましたが、6つの遺伝子をセットで 考える ことができ、われわれは2セットもっていることになります。親から子へそのうちの1セットが遺伝するわけですから、子供は父親とは半分、母親とも半分は一致し ますが、半分は異なります。ただし、兄弟姉妹間をみると、セットの組み合わせは4通りしかありませんので、1/4の確率で一致することになります。
一般に、遺伝子は、たとえばABO血液型の遺伝子にはA型、B型、O型の遺伝子の多型があり、アルコールの分解に関与する遺伝子にも、酒に強い、弱いに関 与 する多型があるというように、数個の多型がみられるのは普通のことです。しかし、MHCの多型は、それらに比べると圧倒的に種類が多いのが特徴です。当然のこ とながら、それには理由があります。
これまで、MHCのことを、単純にタンパク質の断片、ペプチドを乗せるお皿のように表現してきました。しかし、MHCというタンパク質分子とペプチドとい う 分子が結合するには、やはり両者の間に鍵と鍵穴の関係があって適合することが必要です。ただし、ペプチドという分子は小さいため、柔軟性があり、MHCの鍵穴に 形を合わせることがある程度は可能なため、1種類のペプチドしか結合できないというわけではありません。また、一つのタンパク質からは複数のペプチドができま すので、そのうちのどれかがMHCと結合する可能性はあります。しかし、それでも、どのペプチドも結合できないという状況が生じることはあり得ます。となる と、 ウ イルスなどの外来微生物は多種多様ですから、多種類のMHCをそれぞれの個体が保有し、どんなウイルスにも対応できるようにしたいわけです。しかし実際には、 こうしてクラスI、クラスIIそれぞれ6種類ずつ保有しているだけです。本当にこれで十分だろうか、という心配になります。一方、1つの個体が保有するMHC の種類を増やしていくと、図25.2のT細胞受容体の選別で、負の選択を受ける確率がどんどん増えていくことになります。負の選択では、[MHC+自己ペプチ ド] のどれかに強く結合するものが排除されるわけですから、MHCの種類が増えれば、それらのどれかに強く結合する可能性は増え、現状でも正の選択を通過するのは2%程度です ので、それよりもかなり合格率は低くなってしまいます。したがって、むやみにMHCの種類を増やすことはできなかったと考えられます。
それでは、生命系はどういう対応をとったのでしょうか。実は、集団で多様性を維持しようとしたと考えられます。こういうと設計者がいるような表現で誤解を 与 えることになりますが、要するに、MHCが多型化し、一方で1つの個体がもつMHCの種類の数に制約がかかるという進化的な圧力のなかで、自然と集団の中に多 型が維持される構造になり、現在に至っているということになります。ただ、このためには、「性」という、遺伝子の組み合わせをシャッフルしてくれる仕組みがあ る という点が非常に重要となってきます。この点については、本ページ最後の【付録】の「赤の女王仮説」を読んでください。
免疫系では、B細胞受容体(抗体)とT細胞受容体は、1つの個体内で、遺伝子の再構成という手段を使い、膨大な多様性を生み出し、多種多様な外来微生物に 対 抗しています。一方、MHCは、1つの個体内ではわずかな多様性しかもつことができず、集団で多様性を維持してきました。それは、違う視点から見れば、あるウ イルスが流行したとき、もっているMHCの型によって症状の軽い/重いに差が出てくる可能性も示唆しています。どの型のMHCが多いとか、少ないといった型の 分 布には、民族による違いがあることが知られています。そうしたことから、新型コロナウイルスの流行で、発症率に民族による違いが見られましたが、背景にそうした 原因があるのではと論じる研究者もありました。
突然ですが、話題が変わります。1980年代、受精後、細胞分裂が進み、分化していく過程を調べるため、受精後3~4日の発生途上の胚の神経管の一部を、 ウ ズラからニワトリに移植する実験が行われました。こうした異種の細胞が混合されて1つの個体が形成されているものをキメラとよびます。この命名は、ギリシア神 話に登場するライオンの頭、ヤギの胴体、毒蛇の尻尾をもつ怪獣に由来します。しかし、この実験はそうした怪獣を生み出すことを意図としたものではなく、ウズラ とニワトリの細胞が見分けやすいため、移植したウズラの細胞がどのように器官を形成していくのかを追跡することが目的でした。実験は成功し、全体はニワトリの ようですが、移植部分に皮膚の色素細胞に分化する細胞が含まれたため、外見上はっきりと区別できるウズラの黒い羽が生えてきました(図25.4)。

図25.4.胚の神経管の一部をウズラからニワトリに移植する実験(左)。
写真は、左が実験が成功して生まれたニワトリとウズラのキメラ。白い羽はニワトリ、
黒い羽はウズラに由来する細胞からできています。一方、右の写真では、拒絶反応で
ウズラ由来の羽が垂れ下がっています。(写真は絹谷政江氏によるもの)
このキメラたちは、羽を動かし、摂食し、正常に成長しましたが、生後3週間から2ヶ月もすると、まず羽が麻痺して垂れ下がり、歩行も摂食も出来なくなり、 や がて全身が麻痺し、衰弱して死んでしまいました。
その後、孵卵2~3日目の中脳胞(やがて脳を形成する部分)の一部をウズラからニワトリに移植する実験も行われました。成長すると、頭にはウズラ様の黒い 毛が生え、菱脳、小脳、中脳、間脳などがウズラ由来のニワトリが出来あがりました。ヒヨコはウズラと同様な行動をとって鳴いていますが、発声器官がニワトリの ものであるため、鳴き声はニワトリと同じ音質だったそうです。しかし、これも生後十数日で死んでしまいました。
何が起こったのでしょうか?
上で述べたように、未熟T細胞は胸腺で選別を受けます。この場合は、いずれも、胸腺はニワトリのものです。したがって、ニワトリのMHCとニワトリのタン パ ク質の断片の複合体に強く結合するT細胞受容体をもつT細胞は負の選択を受け、死滅し、正の選択を受けたT細胞だけが全身へと広がっていきます。したがって、 ニ ワトリの細胞からなる部分ではもちろん問題ないのですが、ウズラ由来の細胞からなる部分では、MHCがニワトリとウズラでは異なり、タンパク質断片も異なるわ けですから、これに強く結合するT細胞受容体が存在する可能性は十分残されています。そして実際に強く結合すると、免疫系としてはこれを非自己と判断し、排除 しようとします。こうして、ウズラ由来の細胞は攻撃され、最終的には個体の死へとつながったと考えられます。
上記のウズラとニワトリの話は種が違うことがポイントのように思いますが、同じヒトどうしでも、臓器移植をしたときの拒否反応がまさにこれに相当します。
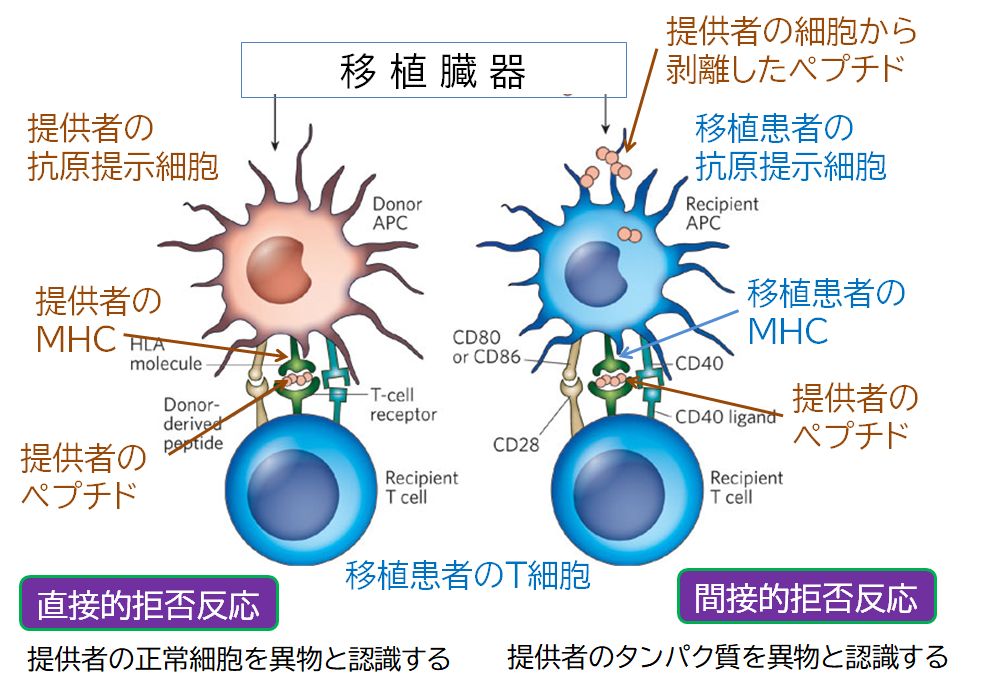
たとえば、提供された臓器の中に提供者の樹状細胞が含まれ、しかもMHCの型が異なるとすると、移植患者のT細胞にあるT細胞受容体は、提供者の樹状細胞 が提示する [MHC+自己ペプチド]複合体は自分のものと異なりますから、強く結合する可能性があります。そうすると、提供者にとっては自己の証である[MHC+自己ペプチド]複合 体であっても、移植患者にとっては、その移植された臓器の細胞上に提示された複合体を非自己の目印と判断し、攻撃することになります。MHCの多様性が臓器移 植では災いとなっています。それはまた、臓器移植を行うとき、MHCの型(いわゆる白血球の型)が一致することが必要と言われる理由でもあるのです。
また、移植された臓器から剥離した細胞や老廃物には、移植患者とは異なるタンパク質が含まれることがあります(MHCはその代表的なタンパク質でもありま す)。われわれはお互いにDNAを比較すると平均0.1% 異なると言われており、作られるタンパク質にもわずかな違いがあります。そうした異なるタンパク質の断片が、移植患者の樹状細胞によって提示されると、このタンパク質を異 物と認識し、そのタンパク質をもつ移植臓器を攻撃することがあると考えられています。
ちなみに、MHC(Major histocompatibility complex主要組織適合性複合体)の名前の由来についてこれまで話してきませんでしたが、組織適合性とは、臓器移植のとき拒否反応が出る原因物質として同定され、この 型が一致していれば拒否反応が出ない、すなわち適合しているというところから名づけられたものです。しかし、臓器移植は自然界で起こる出来事ではありませんの で、生命科学の用語としては何か不適格な感じがします。現在、ヒトに関してだけは、HLA(Human leukocyte antigen ヒト白血球抗原)という呼び名も使われていますが、どちらを使うかは研究者によります。
自己とは何か。この問いには深遠な思想や哲学が含まれた響きがあります。しかし、免疫学的には、胸腺が自己を決める、となります。免疫というシステムの最 大のポイントは自と他の判別であると最初にお話ししましたが、その仕組みがわかっていただけたでしょうか。
次回が免疫に関する最後の講義となります。これまでの話題のなかでこぼれ落ちた花粉症、ワクチン、免疫記憶、腸管免役のお話をしようと思います。
![]()