DNAがもつ情報に関しては、これまで、タンパク質のアミノ酸配列の情報についてのみ述べてきました。しかし、そうしたアミノ酸配列に対応する領域(コー ド 領域)が全ゲノムに占める割合は1.5%もありません。しかも、少数の例外を除いて、個々のタンパク質をコードするDNA上の領域はエクソンと呼ばれるいくつ かの小さな領域に分かれて存在し、その間にはイントロンとよばれる介在配列が存在しています。こうした領域も含めて遺伝子と呼ぶことが多いですが、それでも全 体の25%程度です。ちなみに、その遺伝子を転写するときには、エクソンとイントロンを含む領域がmRNAに転写された後、イントロンに対応する部分を除去し て一本のmRNAへとつなぎ合わせ、タンパク質へと翻訳されるのです。なぜこんなことをするのかは第II部で話そうと思いますので、ここではこれ以上触れませ んが、DNAの塩基配列は、即タンパク質のアミノ酸配列に対応する、というこれまでの私の説明は、あまりにも単純化されていたということになります。
さらに、そのエクソンやイントロンの以外のDNAの領域には、遺伝子がもつ情報の発現、すなわち転写をコントロールする領域があります。たとえば、プロ モーターと呼 ばれる領域は、見た目は遺伝子と変わらず4種類の塩基が並んでいるだけですが、転写を行うRNAポリメラーゼというタンパク質がその塩基配列を認識し、対象とする 遺伝子の転写開始点の目印と しています。RNAポリメラーゼは、いく種類かの転写因子とよばれるタンパク質と複合体を構成し、その領域に結合して転写を開始します。また、エンハンサーと 呼ばれる領域は、そこにアクチベーターとよばれるタンパク質が結合すると転写量が大幅に増大し、逆に、オペレーターやサイレンサーと呼ばれる領域にリプレッ サーなどのタンパク質が結合すると、転写が抑制されるといったことも行われます。

これらの詳細な振る舞いは複雑なので省略しますが、こうした遺伝子の発現の制御が最初に明らかになった大腸菌のラクトースオペロンは、比較的単純で、より教科
書的な話題でもありますので、ここに紹介したいと思います。
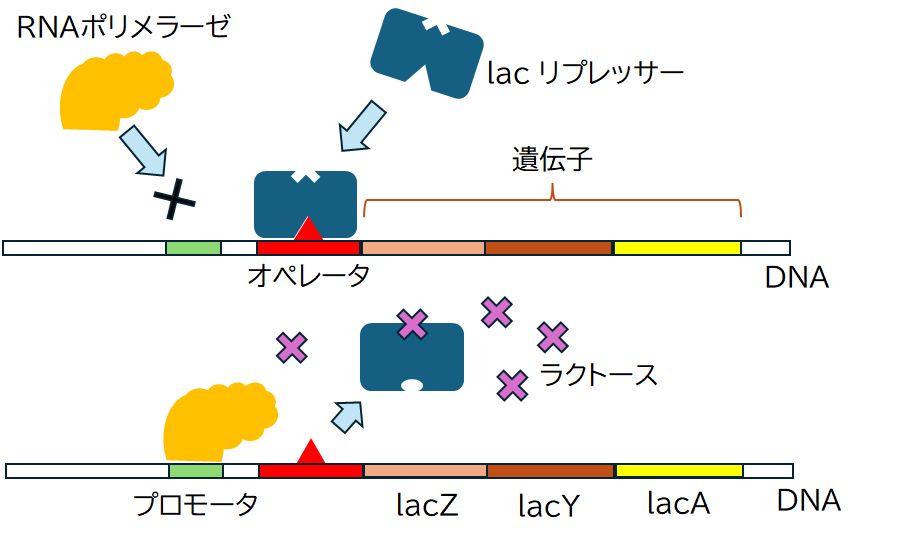
グルコース(ブドウ糖)は大腸菌にとっての主たるエネルギー源です。また、ラクトース(乳糖:グルコースとガラクトースからなる糖)もエネルギー源となるの ですが、利用するためにはラクトース分解酵素を用い、グルコースとガラクトースに分解する必要があります。そのため、グルコースとラクトースがともに存在する 場 合には、グルコースを主として取り込むことがより効率的であるため、ラクトースの取り込みを抑制しています。しかし、グルコースがなくなりガラクトースだけとなる と、取り込みを始め、ラクトース分解酵素の合成も開始します。そこで、そのラクトース分解酵素遺伝子発現のON/OFFをどのように制御しているかをここで見 てみることにします。
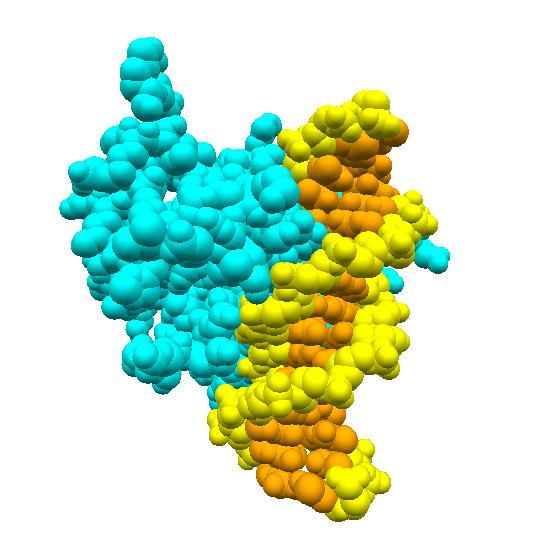
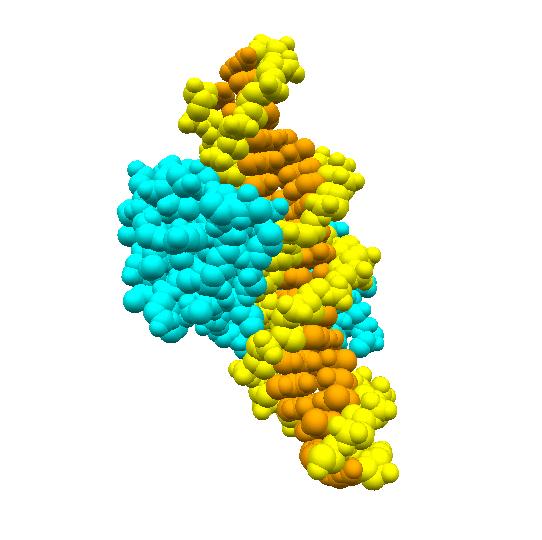
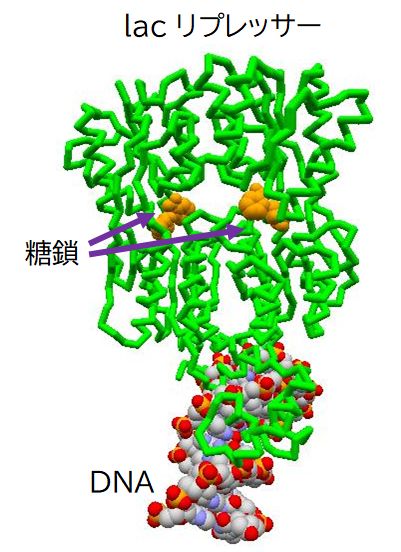
DNAからmRNAへの転写は酵素RNAポリメラーゼが行います。RNAポリメラーゼは、まずラクトース分解酵素遺伝子の前の方にある(上流にあると表現 しま す)プロモーターと呼ばれるDNA上の領域に結合し、その後、ラクトース分解酵素遺伝子へと移動し、転写を行います。ところが、ラクトースがない状況では、 lacリプレッサーと呼ばれるタンパク質が、プロモーターと遺伝子の間にあるオペレーターと呼ばれる領域に結合しています。これが障害となり、RNAポリメ ラーゼはプロモーターに結合することができず、結果としてラクトース分解酵素遺伝子の発現が抑制されます。
ところが、周りにグルコースがなくなり、ラクトースがある場合には、大腸菌はラクトースを取り込むようになります。実は、lacリプレッサーにはラクトース の結合部位があり、そこにラクトースが結合するとlacリプレッサーはその形を少し変え、オペレーターとの間で成り立っていた鍵と鍵穴の関係が崩れ、オペレー ターから離れてしまいます。その結果、RNAポリメラーゼはプロモーターに結合することができるようになり、ラクトース分解酵素遺伝子の転写を行うことができ るようになるのです。
再びラクトースがなくなると、lacリプレッサーに結合していたラクトースも離れ、lacリプレッサーはオペレーター領域に結合できるようになり、ラクトー ス分解酵素遺伝子の転写も休止状態へと戻ります。こうして、転写のON/OFFがラクトースの有無に連動して制御されているのです。
(注)タンパク質が、何かが結合することでその形を変え、その変化が何らかの効果を引き起こすことをアロステリック効果といいます。これについては、このペー
ジの最後の付録を参照してください。
特定の発生過程や生物学的な特性を制御する上で中心的な役割を果たす遺伝子を総称して、マスター遺伝子とよびます。これらの遺伝子から作られるタンパク質は、他 の複数の遺伝子の転写因子として働き、その発現を調節することで特定の生物学的な機能や特性が形成される過程に影響を与えます。
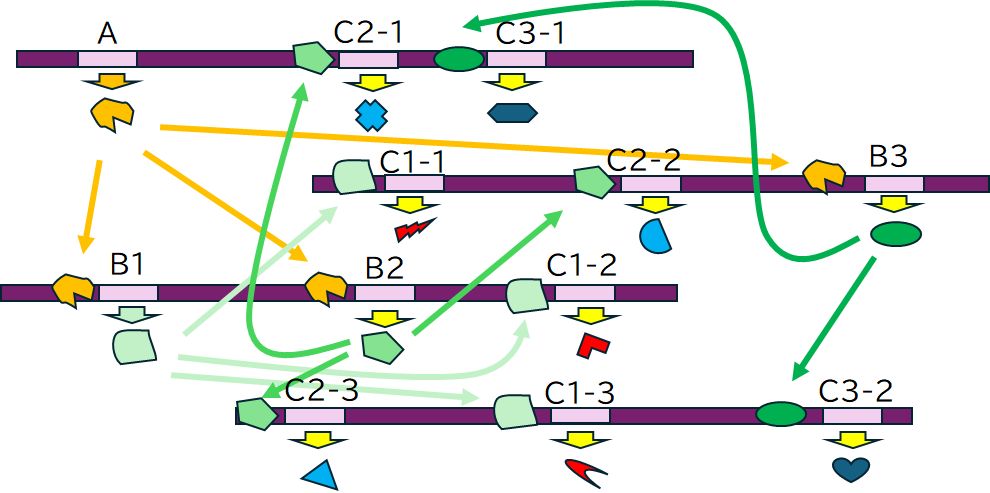
たとえば、多細胞生物の体節形成や器官形成を制御するホメオティック遺伝子がそれにあたります。ホメオティック遺伝子は、多細胞生物の発生において、体節形
成や
器官のパターン形成に重要な役割を果たす一群の遺伝子です。これらの遺伝子は、個々の体節や器官が適切な場所に形成されるように、特定の場所で特定の遺伝子発
現を制御します。
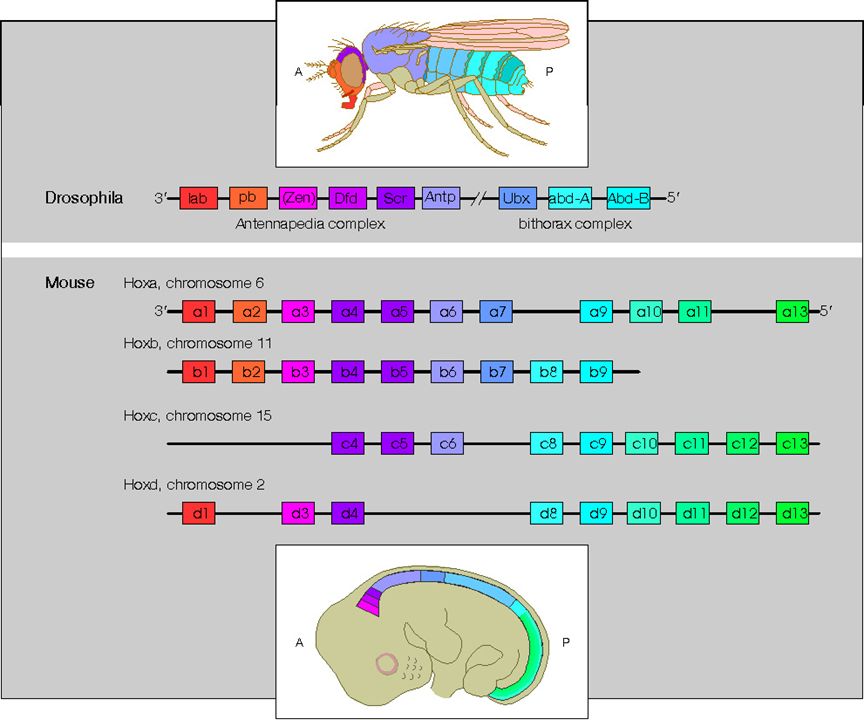
図11.5 ホメオティック遺伝子
(上)ショウジョウバエ、(下)ヒト
ショウジョウバエのホメオティック遺伝子は進化の過程で遺伝子重複を起こし、
哺乳類では異なる染色体上に4個のクラスターを形成している。上図では、相同な遺伝子を
同じ色で示している。ショウジョウバエ、ヒトどちらにおいても、これらの遺伝子が順次発現し
胚の前後軸の形成を誘導する。
図の出典:ホメオボックス @https://bsd.neuroinf.jp/wiki/脳科学辞典
われわれの体はさまざまな種類の細胞から構成されています。しかし、受精卵から細胞分裂を繰り返し、分化していく過程で、その都度すべてのDNAが複製され、受 け渡されています。でも、分化し、専門化した後の細胞では、すべての遺伝子が必要というわけではありません。そして、必要でない遺伝子までが常に活動可能な状 態 でシステムに組み込まれていることは、効率的にも、また誤動作を防ぐ上でも好ましい状況とはいえません。そこで、必要としない遺伝子に鍵をかけ、その活動を抑 制するため、DNAのメチル化ということが行われています。
DNAのメチル化とは、シトシン(C)‐ グアニン(G)と並ぶところのシトシンの一部にメチル基を付加し、DNAを修飾することをいいます。たとえば、メ チル 化がオペレーターなどの転写調節領域で起こると、転写因子との鍵と鍵穴の関係が崩れ、転写因子が結合できなくなるため、遺伝子の発現が抑制されることになるわ けです。
この修飾は、その後の細胞分裂を経ても受け継がれます。DNAのメチル化は、高等生物において、正常な発生と細胞の分化において極めて重要な役割を担っていま す。DNAのメチル化により、細胞が「自分が何者であるか」を記憶できるようになり、安定的にその細胞固有の遺伝子発現パターンを保つことができます。胚発生 の間に〇〇となるようにプログラムされた細胞は、〇〇であるようにシグナルを受け続けなくても、その生物の一生に渡って〇〇であり続けることを保証してくれる のです。
さらに、DNAのメチル化は、宿主のゲノムに取り込まれたウイルスや、その他の有害な要素の遺伝子の発現を抑制することもできます。また、クロマチン構造
の 基礎
を形作るうえでも重要な役割を果たしています。こうして、親から譲り受けた単一不変のDNA配列から多細胞生物に必要な多様な特徴をもった細胞を形成すること
ができるのです。しかし一方で、ひとたびそこに異常が起こると重大な結果をもたらすことにもなります。DNAのメチル化は、ほとんど全ての種類のがんの発達
において極めて重要な役割を果たしていることもわかっています。
![]()
三毛猫 〜 X染色体の不活性化
ラクトース不耐症 vs ラクターゼ活性持続症 〜 牛乳でお腹がゴロゴ ロする
アロステリック効果 〜 タンパク質の形の変化を利用した制御