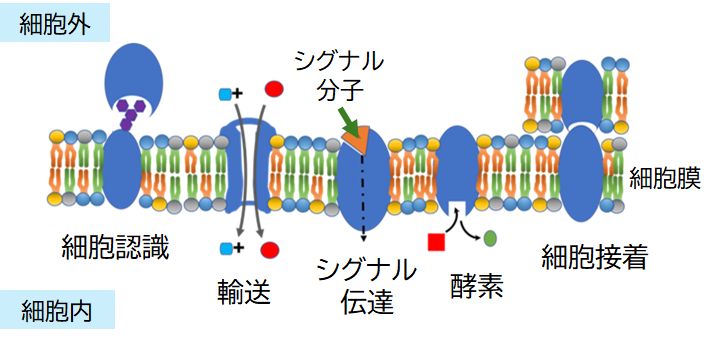
わたしたちは40兆個近くの細胞からなる多細胞生物です。細胞は細胞膜に覆われ、それぞれ独立した世界を構築していますが、孤立しているわけではありませ ん。外部から物質を取り入れ、外部へ物質を排出し、そして他の細胞とコミュニケーションをとっています。それは個体という多細胞からなるシステムを運営してい く 上で欠かすことのできない作業です。そしてこうした役割を担うのが、細胞とその外界とを隔てている細胞膜に埋め込まれたさまざまなタンパク質です(図9.1)。
たとえば、受容体タンパク質(レセプタータンパク質)は、他の細胞から分泌されたシグナル伝達物質(メッセージ物質)が特異的に結合することで、その受容 体 に与えられている特定のシグナルを細胞内に伝達します。多くの場合、シグナル伝達物質は細胞内に入ることはなく、受容体に結合したという情報だけが細胞内に伝 えられます(ただし一部には、細胞内に入り、細胞質または核内に存在する受容体に結合し、情報を伝達するものもあります。性ホルモンはその代表的なもので、本 ページ最 後の【付録】でそれにまつわる話題を取り上げます)。多くのホルモン受容体や神経伝達物質受容体がこれに該当し、この講義で特に注目して取り上げるタ ンパク質です。それは、シグナル伝達物質と受容体タンパク質が鍵と鍵穴の関係にあることが細胞間コミュニケーションを可能にしているからです。
細胞の内外への物質の輸送も避けては通れません。しかし十分に管理して行う必要があります。運搬体タンパク質は、その内部にチャンネルとよばれるトンネル をもち、特定の分 子だけを細胞膜を横切って細胞内へ、あるいは細胞外へと運びます。たとえば、水分子だけを細胞外から細胞内へと輸送するアクアポリンというタンパク質がありま す。チャンネルタンパク質は、トンネルを開閉することで、特定のイオンを細胞内へ、あるいは細胞外へと通過させることができます。主に、神経細胞の興奮(活動 電位の発生)などに関与するもので、ナトリウムチャンネルタンパク質、カリウムチャンネルタンパク質などがあります。こうしたタンパク質も特定の分子のみを通 過させる特異性をもっており、それはこれらタンパク質の立体構造、すなわちトンネルの形によって制御しています。したがって広い意味で鍵と鍵穴の関係で説明で きなくもないのですが、動的な仕組みなので、もう少し適切な表現が必要かもしれません。これらタンパク質については、次回、軽く触れることになります。
その他に、糖鎖を細胞表面に突出させているタンパク質は、細胞の認識マーカーとして機能することが知られています。いま最もホットな研究分野の一つで、そ の機能の詳細はこ れからの課題となっています。ABO血液型の糖鎖も赤血球膜を貫通するグリコフォリンというタンパク質に結合しています。また、細胞が互いに認識・結合し、組 織や器官を形成する上で重要な役割を果たす細胞接着分子とよばれるタンパク質もあります。しかし、これらのタンパク質についてはこの講義では触れられません。
細胞間コミュニケーションの典型的な例がホルモンです。図9.2はそれを模式的に示しています。
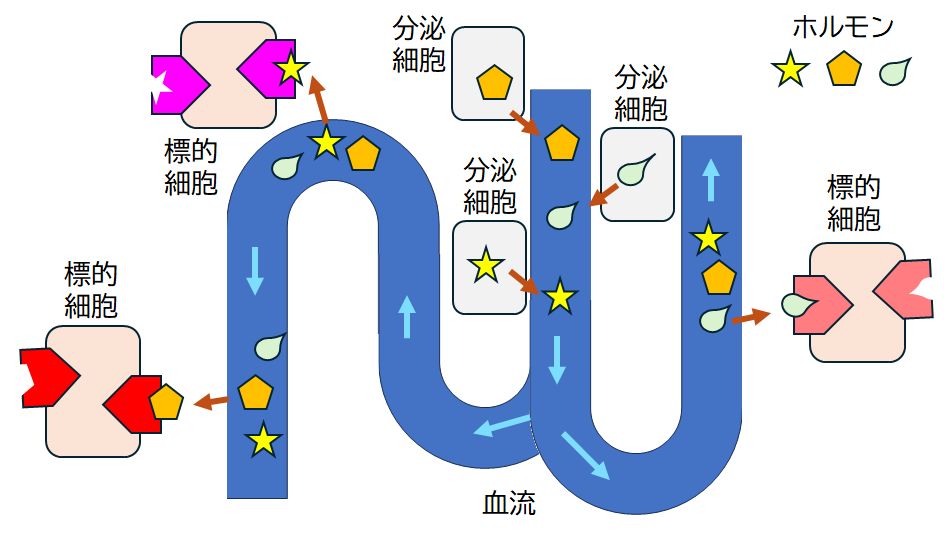
図9.2 ホルモン
ホルモンは、分泌細胞から血液中に分泌され、標的細胞まで血流に乗って移動する。
ホルモンは行先がわかって移動しているわけではなく、鍵と鍵穴の関係にある
受容体タンパク質に出会えば、単にそれに結合するだけである。
内分泌細胞はホルモンを血中に分泌する細胞です。それぞれのホルモンは、それと鍵と鍵穴の関係にある受容体タンパク質にのみ特異的に結合することができる ため、その受容体タンパク質を表面にもつ特定の標的細胞にだけシグナル伝達を行うことができます。 ところで、近年、こうした何らかの“メッセージ”を伝える物質が、内分泌器官とよばれる限られた臓器だけでなく、人体のあらゆる臓器からも放出されていることが次々と発見 されており、その数は数百種類にものぼると言われています。全身の臓器たちは、脳の命令などを介さず、直接メッセージをやりとりし、情報交換しながら、私たち の命や健康を支えていることがわかってきています。
細胞膜から一部を細胞外に露出した受容体タンパク質に伝達物質が結合したという情報は、その受容体タンパク質の細胞内に露出している部分の形の変化を引き 起こすことで他のタンパク 質に伝えられ、さらにそのタンパク質が他のタンパク質へといった一連の反応によって、伝言ゲームのごとく細胞内を駆け巡ります。そして、ある情報は核内のDNAに伝えら れ、ある特定の遺伝子を発現させ、それに対応するタンパ ク質の生合成を促します。またある情報は、外部から細胞内へある物質を取り込んだり、逆に細胞内から細胞外へある物質を排出する作業を開始させます。受容体タ ンパク質へシグナル伝達物質が結合したという情報は、ON/OFFという単純な2値情報ですが、それによって、その細胞にあらかじめ設定されているプログラム に 沿っ た作業が起動され、進行していきます。細胞内ではタンパク質の複雑なネットワークが構築されており、高度な生理現象を実現することが可能なのです(図 9.3)。
(注) ホルモンの種類には、(1) ペプチドホルモン(アミノ酸がつながったもの。ペプチドとタンパク質の違いは、一般に前者が後者より小さいことであるが、はっきりと した境界があるわけではなく慣習的なもの): 成長を促す成長ホルモンや血糖値をさげるインスリンなど、大部分のホルモンがこれに含まれるが、その他にも、リ ンパ球などに作用するサイトカイ ンもこの仲間にはいる。(2) ステロイドホルモン: 副腎皮質ホルモン、性腺ホルモン、ビタミンD3など。 (3) アミノ酸誘導体: 副腎髄質ホルモン、 アドレナリン、ノルアドレナリン、甲状腺ホルモンなど。 (4) プロスタグランジン があります。これらはまとめて生体内情報物質とも呼ばれています。
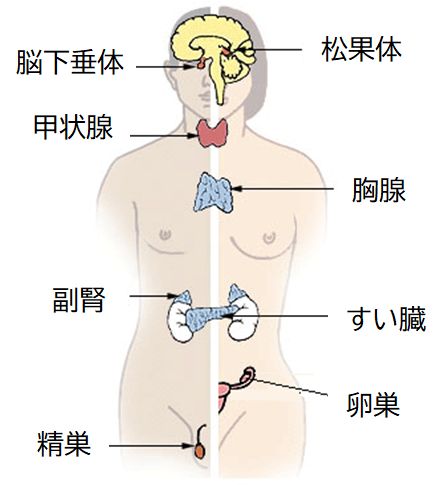
ホルモンをつくる主な内分泌臓器を図9.4に示しました。
インスリンとグルカゴンという2つのホルモンによる血糖値の調節を具体例に、上記の内容を復習してみましょう。
ブドウ糖は重要なエネルギー源で、食事から摂取した後、血液を介して全身の細胞に届けられます。血液中のブドウ糖を血糖とよび、その濃度を血糖値といいいます。とりわけ 脳にとっては使うことのできる唯一のエネルギー源で、その消費量も大きいため、低血糖となって脳が十分なエネルギーを得ることができず機能低下に陥ると昏睡状 態になり、死に至ることもあります。一方、高血糖状態が長期間にわたり続くのは、いわゆる糖尿病です。神経や血管を傷つけ、さまざまな合併症を併発することに なります。したがって、血糖値をコントロールして、ある範囲内に収まるように保つことが必要となります。
血糖値調節の中枢として働くのは間脳視床下部で、ここで血糖値を感知しています。そして血糖値が高くなると、副交感神経を通してすい臓を刺激し、インスリ ンを分泌させま す。インスリンは血流にのって肝臓の細胞に到達すると、その表面にあるインスリン受容体に結合します。インスリン受容体はインスリンが結合したことを細胞内に 伝達すると、血中のブドウ糖を細胞内に取り込み、グリコーゲンを合成して蓄積するという一連のプロセスが起動されます。一方、血糖値が低くなると、今度は交感 神経を通してすい臓からグルカゴンを分泌させます。グルカゴンも血流にのり肝臓の細胞表面にあるグルカゴン受容体に結合します。すると、肝細胞は蓄えていたグ リコーゲンを分解してブドウ糖を生成し、血中へと放出します。
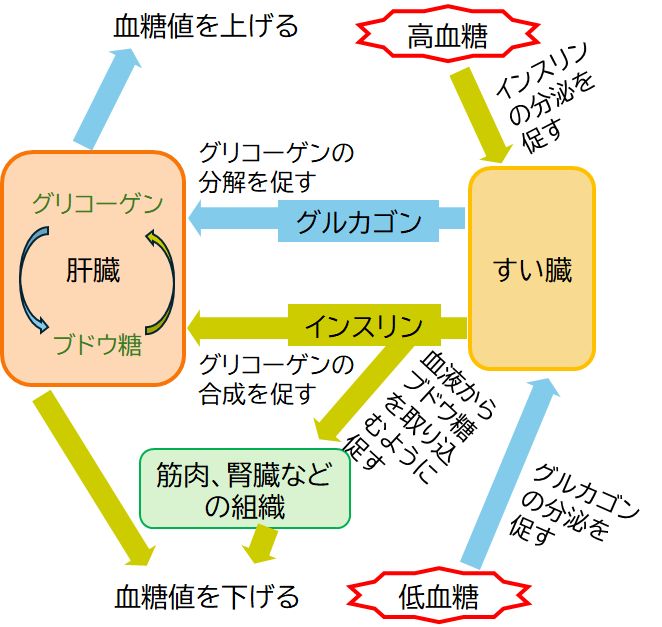
このように血糖値が上がれば下げる方向に、下がれば上げる方向に働くように作られたシステムは、一般に負のフィードバック機構をもつシステムといいます。 もともとワットが蒸気機関を発明したとき、ピストンが速く運動しすぎると蒸気の供給を弱め、遅くなりすぎると蒸気の量を多くして一定の速度で動くように考案し た調速機が起源の工学的な用語ですが、血糖値に限らず、体温や血圧など恒常性を保つ仕組みとして生命システムでも採用されているとても重要な機構です。
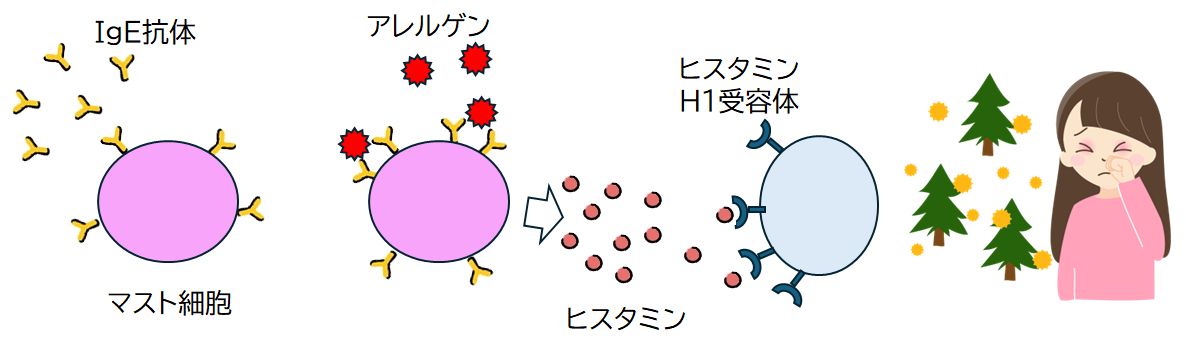
もう一つ身近な例として、ヒスタミンとヒスタミン受容体を取り上げます。 ヒスタミンは肥満細胞と好塩基性白血球において高濃度に蓄えられていますが、細菌などが侵入してきたとき、炎症を惹起するシグナル伝達物質として一過的に放出されます。以 前お話ししたように、炎症は外傷や感染症に対して免疫系が示す重要な生体反応です。また、胃腸にあるエンテロクロマフィン様細胞にも貯蔵されており、摂食によ る胃酸分泌に関与しています。一方、その受け手側のタンパク質、ヒスタミン受容体には、H1、H2、H3、H4の4種類の存在が知られています。とりわけ H1、H2については、H1は花粉症などのアレルギーに関与していること、H2は胃酸過多による胃痛に関与していることなどから身近な存在でもあります。
ヒスタミンH1受容体は、平滑筋、血管内皮細胞、中枢神経系に存在します。ヒスタミンは、H1受容体を介して気管支、胃、腸管など多くの平滑筋を収縮させます。例えば、 ア レルギーによって呼吸困難が起こるのは、ヒスタミンが多量に放出されて気管支の平滑筋にあるH1受容体に結合し、気管支が収縮するからです。しかし本来、これ らの症状は感染に対する生体の防御反応であり、それが過度に反応したものと考えられます。また、末梢神経系のヒスタミンH1受容体は、ヒスタミンの結合によっ て蕁麻疹などのかゆみを引き起こします(これもまた、異常が起きている場所を知らせる生体防御反応の一つと考えられます)。これに対して中枢神経系にもヒスタ ミンH1受容体は広く分布しています。ここではヒスタミンは神経伝達物質としての機能をもち、ヒスタミンの受容体への結合は、脳を覚醒させるというまったく異 な る反応を引き起こします。
一方、ヒスタミンH2受容体は胃壁細胞、平滑筋、リンパ球、中枢神経系に分布しています。たとえば、胃壁細胞にあるヒスタミンH2受容体へのヒスタミンの結合は強力な胃 酸分泌促進作用を示します。気道や血管の平滑筋弛緩、免疫細胞に対する増殖抑制などの作用も報告されています。
ヒスタミンのようなシグナル伝達物質は、ある事態が起こると一過性で放出され、近傍の細胞の受容体に結合し、その事態が起こったことを受容体をもつ特定の細胞に知らせま す。その事態に対応する行動を起こすのはその受容体をもつ細胞の仕事であり、シグナル伝達物質であるヒスタミンはその作業に直接参加することはありません。し たがって、同じシグナル伝達物質が放出されても、放出する側、受け取る側の細胞によって起こる現象は異なるわけです。
第7回の「創薬」の講義では、酵素を標的とする薬の話をしました。実は、受容体タンパク質を標的とする薬もたくさんあります。これには、薬が細胞の受容体に結合し、そ の細胞がもつ働きを促進するようなタイプのもの(受容体作動薬)と、本来の基質の結合を遮断することでその細胞のもつ働きを抑制するタイプのもの(受容体遮断 薬)があります(図9.6)。 ここでは、上で述べたヒスタミンH1受容体とH2受容体の遮断薬の話題をとりあげます。H1受容体とH2受容体はともにヒスタミン受容体ですが、ヒスタミンの結合する向き が異なり、鍵穴の形が異なります。したがって、ヒスタミン以外の薬物では、H1受容体に結合するものが必ずしもH2受容体に結合するとはかぎりません。実際、 以下で述べるH2ブロッカー胃腸薬はH2受容体のみに、抗ヒスタミン薬はH1受容体のみに結合します。
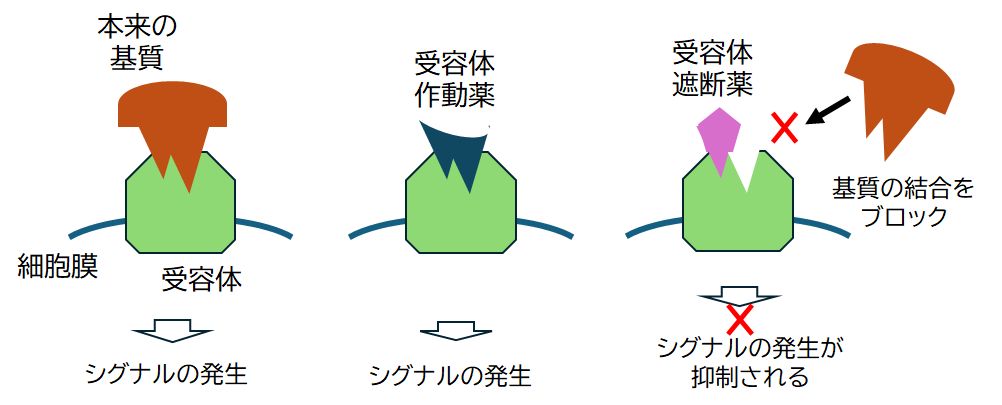
図9.6 受容体タンパク質を標的とする薬
受容体作動薬と受容体遮断薬がある
(1) H2ブロッカー胃腸薬
胃酸は、おいしそうな匂いを嗅いだときなどに脳の指令で副交感神経を通して分泌されるアセチルコリン、胃に食べ物が入ると分泌されるガストリン、胃を常に酸
性に保つため
に分泌されるヒスタミンの3つのシグナル伝達物質によってコントロールされています。なかでもヒスタミンは、胃酸分泌のほとんどケースに関与していると考えられ
ています。しかし、精神疲労や過度なストレスなどさまざまな理由により胃酸の分泌が過多となると、胃粘膜を傷つけ、胃痛の原因となります。さらには、十二指腸
潰 瘍、胃潰瘍、逆流性食道炎なども引き起こします。
そこで、胃壁細胞に存在するヒスタミンH2受容体に結合して、ヒスタミンの結合をブロックするH2ブロッカー胃腸薬がその治療に使われています(図9.7)。
(2) 抗ヒスタミン薬
花粉などのアレルギーの原因物質が鼻に入ると、鼻の粘膜ではアレルギー症状の原因物質であるヒスタミンが作られて体内に放出されます。このヒス
タミンが同じく鼻の粘膜にあ
るH1受容体と結合すると、くしゃみや鼻水といったアレルギー症状(実際には、外部からの異物を排出しようとする生体反応)を引き起こします(図9.8)。そこで、H1受
容体の鍵穴をヒスタミンが結合する前に塞いでしまい、ヒスタミンが放出されても受容体と結合できなくすることで、アレルギー症状の発現を抑えるのが抗ヒスタミ
ン薬です。
その一方で、抗ヒスタミン薬は脳の中にも移行し、脳内にあるH1受容体もブロックしてしまいます。脳内でのヒスタミンの神経伝達物質としての役割は、アレルギー症状の発 現とは無関係で、集中力・判断力・作業能率や覚醒の維持に関与していることがわかっています。そこに抗ヒスタミン薬が入ると、脳内のヒスタミンの働きも妨げら れてしまうのです。その結果、ヒスタミンが脳の中でも働くことができずに、ヒトの活動性が抑えられる、つまり鈍脳を起こす原因となります。これが抗ヒスタミン 薬による眠気の副作用です。
もともと脳には、血液脳関門という機構があって、血液と脳の組織液との間の物質交換を制限するように働いているのですが、第1世代抗ヒスタミン薬はそこをすり抜けていま した。しかし、第2世代の抗ヒスタミン薬では、その脳関門を通りにくくするように改善され、眠くなりにくくなっています。
![]()
環境ホルモン 〜 環境に放出された人工的な化 学物質がホルモンに似ているために・・・
フィードバック機構 〜 ホメオスタシス;生命の恒常性を保つために・・・