前回は少々横道にそれて創薬のお話をしましたが、今回も、薬といえばやはりウイルスのことが気になるというわけで、もうしばらく寄り道 をしてみたいと思います。 ただし、話のキーポイントは、これまでと同様、タンパク質と基質の「鍵と鍵穴の関係」です。
ウイルスは生物か、生物ではないかがよく議論になります。この講義の最初に生命を定義することは難しいという話をしましたが、所詮、定義の問題ですから、 生 命だとしたい人はウイルスも含まれるように生命を定義すればいいし、外したい人はそのように定義すればいいでしょう。でも、それが議論になるということは、ウ イルスがそれ以外の生物と比較するとかなり特殊な存在であるということです。核酸とタンパク質からなるシステムという意味ではこれまで話ししてきた生物の 範疇にはいりますが、以下の点ではウイルス以外の生物と大きく異なります。
・ 細胞質などはもたず、タンパク質と核酸からなる粒子である(結晶化することもできます)。
・ 遺伝情報物質としてDNAかRNA、どちらか一方だけしかもたない。
・ 自身の代謝系はなく、宿主細胞の代謝系を利用する。 そのため、単独では増殖できず、他の生物(宿主)の細胞内に侵入し、その生物のシステムを借用して増殖する。ただし、宿主細胞の代謝系で働く自前のタンパク質を いくつかもつ。(このことは、何らかの細胞に侵入しないかぎりは単なる物質に近く、侵入して初めて生命系に何らかの作用をもたらすことを意味します。ウイルス による感染症を考えるとき、このことは重要な示唆を与えてくれます)
・ ゲノムサイズは万から数十万塩基程度で、他の生物と比べ格段に小さい(例えば、大腸菌で460万塩基対、ヒトでは30億塩基対からなる)
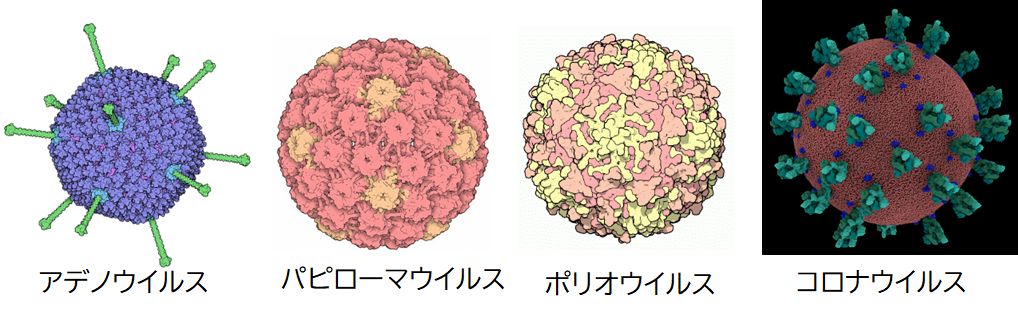
図8.1にみるように、形状は球状のものが一般的ですが、棒状のものなどもあります。図8.1で彩色されている表面の凹凸もしくは突起はタンパク質からで きており、ウイルスの種類を特徴づけています。特に、抗原性を決める重要な要素の一つであるため、感染症の観点からは特に注目するポイントとなります。
ウイルスは、その内部に埋め込まれた遺伝情報物質がDNAかRNAかによって分類することができます。DNAウイルスには、アデノウイル ス、ヘルペスウイルス、パピローマウイルスなどがあります。RNAウイルスには、風疹ウイルス、ノロウイルス、インフルエンザウイルス、はしかウイルス、コロ ナウイルス、HIVウイルス、B型肝炎ウイルスなどがあります。一般に、RNAウイルスの方が変異が速く、一度できた抗体が効力を失う確率が高いため、われわ れにとって厄介な存在となっています(第4回 講義 4.1 DNAとRNAの(注)も参照してください)。
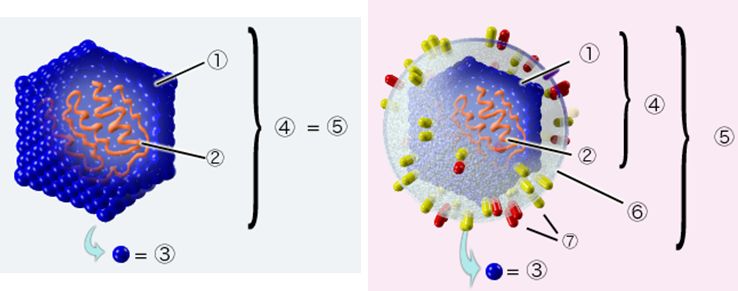
ウイルスはその基本構造で分類することもできます。どのウイルスも、遺伝子である核酸(DNAかRNA)をタンパク質の殻(カプシド)が包み込んだ構造か らできています。ウイルスによっては、そのカプシドのさらに外側に脂質と糖タンパク質からなる被膜(エンベロープ)が存在するものもあります(図8.2)。エ ンベ ロープの主成分である脂質は、アルコールや界面活性剤(洗剤)で破壊されやすいため、エンベロープウイルスであるインフルエンザウイルス、新型コロナウイルスに はアルコール消毒や石鹸による手洗いが除菌対策として有効となります。一方、エンベロープをもたないノロウイルスなどでは、アルコールや界面活性剤は効果があ り ません。これらウイルスには次亜塩素酸ナトリウムが有効であることが知られています。
ウイルスの増殖(ライフサイクル)は一般に以下のステップで行われます。
宿主細胞表面への吸着 → 細胞内への侵入 → 脱殻 (殻内部から核酸の遊離)
→ ウイルスを構成するタンパク質の合成(ウイルス遺伝子の発現)
→ 合成されたタンパク質の集合・組立て → 感染細胞からの放出
ここで、この講義にとって重要な点は、最初の宿主細胞表面への吸着から細胞内への侵入において、ウイルス表面のタンパク質(図8.1の彩色した部分)と宿主細胞表面にあ る 何ら かの分子(受容体分子)とが鍵と鍵穴の関係にあることです。一般に、生物の細胞表面にはさまざまな分子が存在します。それは生物種によっても異なりますし、同じ 個体内でも、器官・組織によって異なります。したがって、ウイルスはどの細胞にも侵入できるわけではなく、鍵と鍵穴の関係にある受容体分子を表面にもつ細胞に のみ侵入できるわけです (もちろん、その分子はウイルスが侵入するための受容体として存在しているわけではありません。宿主となる生物にとって何らかの機能を 果たしている分子なのですが、偶然、不幸にもウイルスのもつタンパク質と鍵と鍵穴の関係になっていたということです) 。一般に、ウイルスは、ある生物種にのみ感 染するという種の壁(宿主特異性)がありますが、こうした理由によります。鳥インフルエンザがヒトには感染しないのもそのためです。しかし変異を起こし、ウイ ルス表面にあるタンパク質の形が変わってヒトの受容体に結合できるようになると、ヒトに感染することが可能となり、新型インフルエンザの誕生となるわけです。
新型コロナウイルスが猛威を振るっていたころ、「感染」という言葉に何かもやもやした感じをもって いました。ウイルスに感染したとは、体内にウイルスが入ったときか、ウイルスが細胞に侵入したときか、症状が出たとき、つまり発症したときか、がなんとなく はっきりしないまま使われることが多くあったからです。上で述べたように、ウイルスは自分自身で増殖することはできず、宿主の細胞に侵入しない限り何もできま せん。したがって、宿主の細胞に侵入したときが感染、というのが正確な定義ということでしょうか。一方、ワクチンを接種したり、実際に感染したりして抗体がで きたとしても、体内にウイルスがはいってくることはあります。しかし抗体があることで、感染する前に排除されたり、感染しても初期段階で消滅させられ発症まで 至らない、そうした確率が非常に高くなることは確かですが、ワクチンをしていれば感染しないという表現は厳密に考えると微妙です。また、初めての感染でも、自 然免疫、獲得免疫(いずれ話題にする予定です)によってウイルスが排除され、発症まで至らない場合もあります。発症しなければ普通感染したとは言わないでしょ う。などと考えていると、「感染」という言葉の使い方が実にあいまいで、心にもやもやが残ってしまうのです。
インフルエンザ・ウイルスを例に、図8.3を見ながらもう少し具体的に上記のことを見ていきましょう。
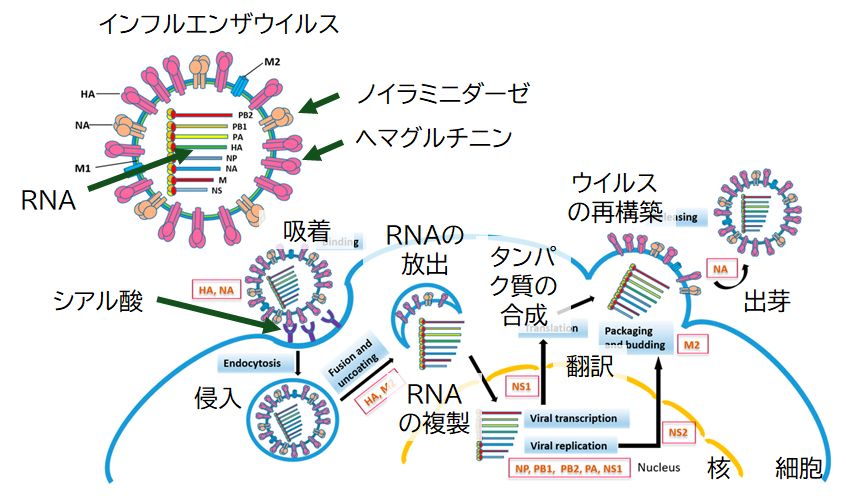
インフルエンザ・ウイルスは体内に入ると、① その表面にあるヘマグルチニンというタンパク質が、人の細胞表面にあるシアル酸(糖の一種)に結合(吸着)することで、② 人の細胞の内側に侵入することができます。(すなわち、ウイルスがもつヘマグルチニンと宿主細胞にあるシアル酸が鍵と鍵穴の関係にあり、特異的に結合します)③ ウイルスは、その遺伝情報物質である RNAと内部にもっていた自前のタンパク質を宿主細胞の核へと放出します。④ 放出された タンパク質によってRNAが複製されます。⑤ 複製されたRNAはmRNAとして働き、宿主細胞のシステムを利用して、ウイルス自身に必要なタンパク質を合成 します。⑥ 増殖したウイルスがもつためのRNAも複製し、⑦ 最終的に、複製されたRNAと合成されたタンパク質を組み立て、新たなウイルスとして宿主細胞を飛び出していきます。大量の子ウイルスが飛び出していくことで宿主細胞は破 壊されたり、障害を受け、その器官に何らかの症状が出ることになります。
インフルエンザ・ウイルスが宿主細胞を飛び出すとき、エンベロープ部分には宿主細胞の細胞膜が利用されます。そのためウイルス表面には宿主細胞にあったシアル酸が存在し ます。このことがウイルスにとっては困ったことを引き起こします。飛び出したばかりのウイルスが他のウイルス表面にあるヘマグルチニンと結合する可能性がある からです。また、宿主細胞表面のシアル酸と再び結合す る可能性もあります。もしそうなると身動きがとれなくなるため、ウイルスは、シアル酸とヘマグルチニンの結合を切断する機能をもつノイラミニダーゼと呼ばれる酵 素をその表面にもっています。シアル酸を切り離すことで自由に活動することができるようになり、ウイルスは新たな細胞へとさらに感染を広げていくことができる のです。
ちなみに、インフルエンザ・ウイルスは、大きくA型、B型、C型の3つの型に分類され、さらにA型では、ウイルス表面にある2つのタンパク質ヘマグルチニン(H)とノイ ラ ミニ ダーゼ(N)の抗原性の違いによって、HとNに番号をつけて区別する亜型があります。現在、ヘマグルチニン16種類、ノイラミニダーゼ9種類が報告されており、 H1N1~H16N9までの亜型があります。
(注) 抗原性の違いによって、ヘマグルチニンにはH1~H16の亜型とノイラミニダーゼにはN1~N9までの亜型があるとは、亜型ごとにこれらタンパク質のアミノ酸配列 が一部異なり、その立体構造が一部異なることを意味しています。抗体(これもタンパク質です)はヘマグルチニンやノイラミニダーゼに結合するのですが、当然の ことながら「鍵と鍵穴の関係」で結合します。しかし、H1と結合できた抗体が結合できないヘマグルチニンがあるとすると、それは、RNAが変異し、アミノ酸配 列が変化し、立体構造が変化したからで、新たな亜型として新たな番号が振られるわけです。立体構造の違いを調べるのは結構面倒なので、実際には既存の抗体が結 合する かどうかで抗原性の違いを判断しますが、この講義の流れでいえば、それは立体構造の違い、すなわち鍵と鍵穴の関係性の違いで区別しているということになりま す。
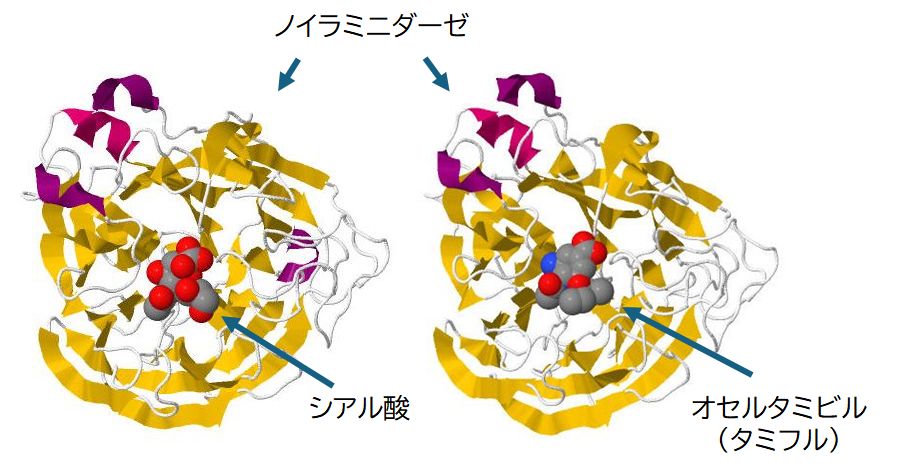
タミフルはインフルエンザ治療薬のいまや定番となりましたが、この薬は、ウイルスのもつ酵素ノイラミニダーゼを標的とし、これを阻害するよ うに設計されたものです。したがって、宿主細胞から飛び出してきたウイルスがシアル酸と再結合して身動きが取れなくなったその状態のままにウイルスを留めてお こうというもので、ウイルスを殺す機能はなく、その増殖を抑制するものです。これらの薬は感染初期の増殖がまだ進行していないときに服用するのが効果的とされ るのはそのためです。 タミフルはノイラミニダーゼの基質であるシアル酸に類似し、しかし、シアル酸より容易に、より強くノイラミニダーゼに結合するようにデザインされた化合物です。
ウイルスのなかに、レトロウイルスとよばれる一群があります。エイズのHIVウイルス、成人T細胞白血病ウイルスなどがこれに属します。RNAウイルスで すが、インフルエンザ・ウイルスなどのその他のRNAウイルスとは大きく異なる特徴をもち、興味深いので紹介しておきます。
ウイルスが宿主細胞表面にある受容体となる分子に結合し、細胞内に侵入、RNAを放出するところまではインフルエンザ・ウイルスなどと共通です。ところが、レトロウイル スはここで、自らがもっていた逆転写酵素によって、RNAからDNAへの逆転写を行う点が決定的に異なります。第4回で説明したDNA→RNAという転写とは 逆の情報の流れ方です。出来上がったDNAは宿主細胞の核へと移動し、そこにある宿主のDNAへ組み込まれます。
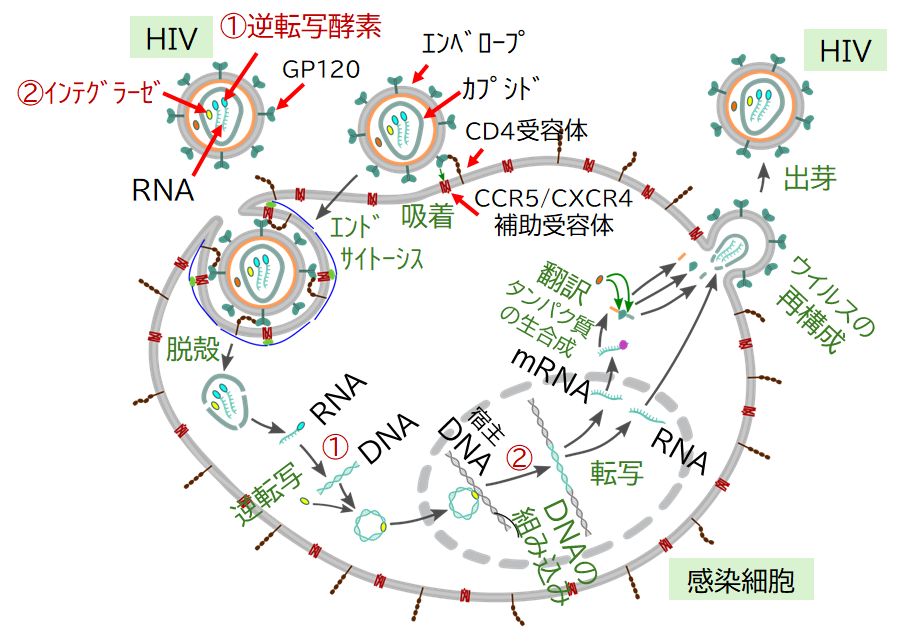
図8.5の一番下に注目してください。ウイルスのRNAが自前の逆転写酵素によってDNAに逆転写され、それが、灰色で描かれた宿主のDNAに組み込まれ る様子が描かれています。塩基配列が違うだけで、化学物質としてのDNAという点では逆転写でできたウイルスDNAも宿主DNAも何の違いもありません。 その後、そのまま何も起こらない場合もありますが、何らかのきっかけでこの組み込まれたDNAからmRNAに転写され、タンパク質への翻訳が始まります。合成 されるタンパク質はもちろんウイルスのタンパク質です。それらのタンパク質と転写されたRNAを組み上げればレトロウイルスが完成するという仕組みです。こう して大量のウイルスが1つの細胞内で作られ、宿主細胞から飛び出していきます。細胞は破壊されたり、損傷をうけ、その結果宿主に何らかの病状を引き起こすこと になります。ちな みにエイズのHIVは、ヒトのある種の免疫細胞にある特殊な分子を受容体として感染するため、その免疫細胞が破壊されることで免疫不全の症状があらわれます。
レトロウイルスが宿主細胞のDNAのどこに組み込まれるかは決まっていません。したがって、遺伝子領域に割り込んでくることもあります。そのときには、その遺伝子から作 られるはずのタンパク質が作られなくなり、遺伝子によってはそれが原因でがんなどを引き起こすことがあることも知られています。
(注)HIVがT細胞やマクロファージなどの免疫細胞に入りこむには、細胞表面にあるCCR5とい うタンパク質に結合することが必要です。しかし、ヨーロッパではごく稀に、この CCR5遺伝子が欠失しているため、CCR5タンパク質を細胞表面にもたない人がいます。そして、この欠失をもつ人は HIV に感染しにくい(明らかにウイルスに暴露されているのに発症しない)ことが報告されています。
(注) 自身のゲノムを宿主のDNAに挿入することのできるレトロウイルスの性質を利用して、動物細胞に外来遺伝子を組み込み、その遺伝子からタンパク質を作らせることが できま す。こうした遺伝子の運び屋としての能力をもつウイルスをベクターといいます。遺伝子操作によりウイルスの一部の遺伝子を欠損させて複製および増殖能をもたな いようにしたのち、外来遺伝子を組み込み、対象とする細胞に感染させます。遺伝子疾患を持つ患者の細胞に、その疾患の原因である変異遺伝子の正常型を直接導入 することで治療を行う遺伝子治療も研究が進んでいます。
![]()