前回、進化をいかに捉えるかという観点から、特に「自然選択」を巡るいくつかの論点についてご紹介しました。今回の講義は、それに答えるべくいくつかの話 題 を取り上げますが、あくまでも一つの考え方を紹介するだけで網羅的な解説はできませんので、こうした話題を基礎に自分なりにいろいろ調べたり、考えたりしてい ただければ幸いです。
前回の講義において、自然選択において何が選択されるかという問いに対して、集団選択説、個体選択説、遺伝子選択説があることを紹介しました。この問題に ついて考えるために、あるたとえ話から始めたいと思います。
チームで行うスポーツ、たとえば野球とか、サッカーとか、ラグビーを思い浮かべてください。いま最強のチームを作るべく、多数いる候補選手から一流選手を 選 出したいのですが、選手のプレーを観察してもその能力を見抜く能力がまったくないとします。こうした場合、どのようにしたら一流の選手を選べるでしょうか?そ んな問題を考えてみます。
そこでまず、多数いる候補選手をランダムにグループ分けしてチームを作り、それらのチームで総当たりのリーグ戦を行うことにします。一試合一試合の勝ち負 け には運もありますが、試合数をできるだけ多くし、その勝率から最強のチームを決めることはそれなりに合理的な方法といえるでしょう。しかし、その優勝チームが 求めている最強のチームかと問われれば疑問符が付きます。各チームの選手はランダムに振り分けられていますから、その優勝チームの中に優れた選手が含まれてい ることは確かでしょうが、全員が優れている選手とは限りません。また、個々の選手の能力を見抜く能力はないと仮定しましたので、リーグ戦を観戦する中で、優勝 チーム以外のチームも含めて、優秀な選手をピックアップすることもできません。したがって、これだけでは一流の選手からなるチームを作るという当初の目的を達 成することはできません。
そこでチームをすべて解散し、再び選手たちをランダムにグループ分けし、同じことを繰り返すことにします。しかし、何も新しい要素はありませんから、ふた た び優勝チームが決まったとしても結論は同じです。それでもまた、すべて解散してはランダムにグループ分けしリーグ戦、解散してはランダムにグループ分けしリー グ戦を繰り返すことにします。そうした中で、選手ごとに、属したチームの全勝率を計算します。その勝率が高い選手はチームの勝利に貢献した可能性が高い、すな わ ち優秀な選手といえないでしょうか。属したチーム数が少なく、試合数も少ないとすると、実力がないのに偶然勝ったチームにいた可能性を捨てきれません。しか し、 次から次へとチーム編成が変わり、非常に多くの試合を行った末、高い確率で勝つチームにいたとしたら、それは単なる偶然ではなく、その選手のチームの勝利への 貢献度はかなり高いと評価できるのではないでしょうか。
この話は、個体をチームに、遺伝子を選手にたとえたものです。自然選択の直接の対象は明らかに個体です。しかし、どんな優秀な個体でも寿命があります。子 孫 がその遺伝子を受け継ぐと言えますが、有性の種では必ず配偶者の遺伝子と混合しますので、その個体としての優秀さがそのまますべて子孫に伝わるわけではありま せん。となると、個体が自然選択の対象であるとはどういう意味があるのでしょうか。そんな疑問について考えるためのたとえ話でした。
たとえ話では、試合の勝敗が評価の全てであり、選手に関しては直接評価をしていません。自然選択が個体に対して行われることに対応します。しかし、チーム は あ る期間が過ぎると解体され、新たに編成されます。生物でも配偶者の遺伝子と混合させて、自らの遺伝子の半分を子孫に引き継ぎ、個体はやがて寿命が尽きます。ま たどんな強いチームといえども、その中のすべての選手が優秀というわけではなく、どんな優れた個体といえども、そのすべての遺伝子が優れているわけではありま せ ん。たとえ話のように、まったくランダムにチーム編成はしませんが、配偶者との間で遺伝子のシャッフルが行なわれながら、何世代も何世代も経て、今生きている個 体 がいます。その長い年月を経て、一体何が選択されたのでしょうか。それは自然選択された個体の中を連綿と受け継がれてきた遺伝子ということにならないでしょう か。これが遺伝子選択説の基本的な考え方であり、このたとえ話の趣意です。
進化の理解の難しさの一つは、進化がこうした集団-個体-遺伝子という階層構造のなかで起こる事象であり、直接的な選択は個体レベルで起こるとしても、実 質 的に選ばれているのは遺伝子であるという点にあるといえます。しかも、さらに深く検討するとなると、選手間の相性という問題も考える必要があるでしょう。勝率 の高いチームには必ず優秀な選手が含まれていると述べましたが、飛び抜けた選手がいなくても、チームワークによる勝利もありえます。ある遺伝子の組み合わせ が、 ある環境での個体の適応度を高めることが考えられます。進化の論理構造を組み立てることがいかに難しいかがわかっていただけると同時に、こうした論理を追求す ることがいかに興味深いことであるかも理解していただけたのではないでしょうか。
前節で、自然選択は集団、個体、遺伝子という階層構造の中で、どのレベルに作用するのかを問題として取り上げ、遺伝子に注目しました。しかし個体と集団と の 関係性についても調べる必要があります。また、自然選択にとって環境への適応度が選択の基準となるというときの環境とは何かについても考えてみなければなりま せん。こうした問題を考えるために、動物の行動の進化を論じるときに利用される「タカ・ハトゲーム」を次に紹介しようと思います。
動物の行動には、生得的なもの、いわゆる本能行動がありますが、これは遺伝子によって決まっていると考えられます。遺伝子とは、タンパク質のアミノ酸配列 を 指 定するDNA上の領域ですから、それがどのように生得的行動をコントロールするかは謎ですが、生後の教育で後天的に得られたものではない種特有の行動パターン は、遺伝子によって伝えられたと考えるしかないということになります。そこで、疑念は残りますが、動物には遺伝子によって制御された行動パターンがあると仮定 し て話を進めます。すなわち、これから述べる動物の行動は、その動物の意思とか感情とかに一切関係なく、遺伝子の命令のままの振舞いと捉えます。また、話を簡略 化するために、性のことは考えず、ある個体がもつ遺伝子はその子に必ず伝わるとします。
さて、生得的な動物の行動パターンのなかに、同種の個体どうしがかなり激しい戦いを繰り広げた場合でも、最終的に勝者が敗者に対してとどめを刺すことはな い というものがあります。これは、ノーベル生理学医学賞受賞者ローレンツの代表的な業績の一つで、多くの観察例から、同種の個体を殺すことに強い本能的な抑制が かかっていることが明らかとなっています(例外的な種が身近にいますがそれは横に置いておきましょう)。同種の個体どうしは、餌をめぐって、配偶者をめぐっ て、縄張りをめぐってのライバルであり、殺した方が自らの、そしてその子孫の生存率を上げるという意味において進化的なメリットがあるように思えます。にもか かわらず、同種の個体は殺さないという行動をとる遺伝子が集団に固定されたのはなぜなのでしょうか。
そんなことをしたら種が滅びてしまうから、あるいはそれは「種にとっての善」とローレンツをはじめ多くの動物行動学の研究者は説明してきました。これは集 団 選択説的な考え方です。これに対して、個体選択説や遺伝子選択説を主張する研究者たちは、あくまでも自らの生存率を上げ、その要因である遺伝子を子孫に伝え、 その遺伝子をもつ子孫の生存率をあげることを基本原理とするのが進化の論理であって、同種の他の個体の生存率などどうでもいいはずだと考えます。みんなで仲良 くという利他的な 論理は倫理的には好ましいかもしれないが、進化の論理としては科学的ではないというのです。しかし、そんな利己的な個体の集まりにおいてもなお、同種の他の個 体を殺さないという利他的行動を促す遺伝子が進化の過程で選択されることを説明することが可能なのでしょうか。実は、それを説明するためのもっとも簡単なモデ ルとして提唱されたのが、タカ・ハトゲームなのです。
タカ・ハトゲームでは、生得的行動の遺伝子として次に掲げる2種類の遺伝子、タカ派戦略の遺伝子とハト派戦略の遺伝子が存在し、集団内の各個体はそのどちら かを保有しているものとします。そして、各個体の行動はその遺伝子の戦略に完全に支配されていると仮定します。ここでゲームとか戦略という言葉が出てきたの は、 ここで利用する数学的手法が、経済学で使われているゲームの理論をベースにしたもので、そこでの用語がそのまま使われているからです。
いま、食料や配偶者や縄張りを手に入れるため、同種の2つの個体が対峙しているとします。このとき、タカ派戦略の遺伝子は、自分が傷つくか、相手が逃げだ すま で戦いを挑み続けるよう個体に指示します。一方、ハト派戦略の遺伝子は、まずはそれを欲しいと相手に誇示しろと命じますが、もし相手が戦いを挑んできた場合に はただちに逃げ出すよう指示します。
このモデル設定では、タカ派の個体とハト派の個体が出会うと、タカ派の個体がすべてを得ることになります。したがって、やはり、ライバルとは徹底的に戦う こ と が進化的に有利であり、気弱なハト派に生き延びる術はないように思われます。ところが集団として考えると必ずしもそうとは言えない、というのがこのモデルのポ イントです。
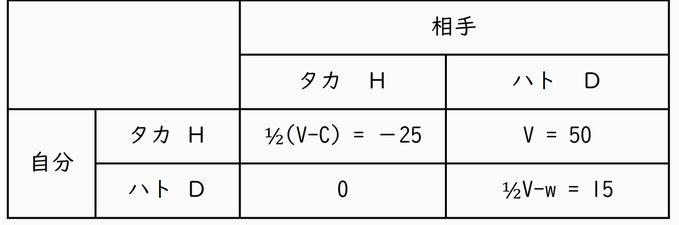
タカ・ハトゲームの利得表
利得: V = 50, C = 100, w = 10
最初の議論ではタカ派とハト派の出会いだけが想定されました。タカ派がすべてのV (= 50) を獲得し、ハト派の利得は0です。しかし、集団として考えれば、タカ派とタカ派の出会いもありますし、ハト派とハト派の出会いもあります。タカ派とタカ派が出会うと徹底的 に 戦いますから、勝つこともあれば負けることもあります。勝てば利得Vを得ますが、負ければ傷ついて適応度を下げることになり、そのコストがC (= 100)で与えられています。勝つ/負けるは5割の確率で起こるとすると、平均の利得は ½(V-C) と設定することができます。一方、ハト派とハト派の出会いでは1/2V と設定していますが、どちらかが身を引く確率が1/2と考えてもいいですし、仲良く半分ずつ分け合うということで1/2V としたと考えてもいいでしょう。w (= 10) が引かれているのは、両者が出会った当初はにらみ合いに時間を費やすため、その時間を他に有効に使えば得られたかもしれない資源の価値を考慮したからで、にらみ合いのコス トといってもいいかもしれません。ただ、ここでの議論ではあまり本質的ではありませんので、有っても無くてもいいものです。
ここからは、ゲームの理論を使って数式を導き、説明することが本来のやり方ですが、ここでは定性的な説明だけをします。詳細を知りたい方は「タカ・ハトゲー ム」をキーワードに自分で調べてみてください。
ゲームの理論を使って解析する上で重要な点は、集団内のハト派の個体数とタカ派の個体数の割合です。ハト派の個体がハト派の個体あるいはタカ派の個体に出 会う確率、そしてタカ派の個体がハト派の個体あるいはタカ派の個体に出会う確率がその割合によって異なり、それによって得られる利得が異なるからです。
そこでまず、集団内の個体が全員ハト派としてみましょう。争いのない平和な集団です。しかし、そこにあるとき突然変異が生まれ、その個体がタカ派となっ た、 あるいは他の集団からタカ派の個体が侵入してきたとします。まわりはすべてハト派ですから、このタカ派の個体は何もかも手にいれることができ、その子孫は急速 に数を増やしていくことになります。
それでは、最終的にタカ派の個体だけの集団になるのでしょうか。そこで今度は集団内の個体がすべてタカ派の場合を考えてみましょう。争いが絶えず、各個体 は 1/2の確率で勝ったり負けたりします。しかも傷つくことのコストの方が得られる利得よりも高いと設定されているため、平均の利得は −25とマイナスの値となっています。そうした中に、やはり突然変異や移住によってハト派の個体が混じると、上の利得表ではハト派の適応度の変化は0ということですが、生 存に必要な最低限の資源は得られるとすると、−25よりは0の方が大きいですから、ハト派の子孫が増えていくことになります。
結局、全員タカ派も、全員ハト派も安定な集団ではないということになります。ということは、ハト派の個体とタカ派の個体それぞれの利得が釣り合う割合があ る ことが予想されます。実際、ゲームの理論による解析から、上の例ではタカ派 7/12、ハト派 5/12で両派の個体の利得が等しくなり、この割合が平衡点となります。
この議論は、一つの個体がときにタカ派的に、ときにハト派的に振舞うというモデルにすることも可能です。いろいろな確率でタカ派的に振舞ったり、ハト派的 に 振舞ったりすると仮定すると、7/12の確率でタカ派的に、5/12の確率でハト派的に振舞う個体からなる集団へと最終的に収束することを意味しています。
また、平衡点はタカ派どうしでの戦いで設定されたコストCの値に大きく依存します。この値が小さければ、すべてタカ派という状態もあり得ますし、大きけれ ば、さらにハト派の割合が大きな集団が平衡点となります。相手を殺してしまうというのは、殺されるというリスクも負うことであり、少なくともこのモデルでは、 タカ派が集団内の圧倒的多数派にはなり得ないことを意味します。
もちろんこのモデルは動物の行動を理解する上ではあまりにも単純なものですから、突っ込みどころはいろいろあると思います。しかし、タカ派の個体とハト派 の 個 体を比べ、タカ派の個体の方が強いというだけでは集団としての振舞いは説明できないという点を理解していただくこと、そしてその集団としての振舞いを、種が滅 びてしまうから徹底的に戦わないという集団選択説の論理を使わなくても、タカ派的振舞いとハト派的振舞いを適度に混合させて振舞うことが安定だから、あるいは 2つのタイプの個体あるいは遺伝子の頻度がある割合で釣り合うからという論理で説明できることを理解していただくことが重要だと思います。
このタカ・ハトモデルは、個体にどのような行動を命令する遺伝子がその生物種の子孫の中に広まっていくかという遺伝子選択説の立場からの説明でした。しか し、「種が滅びてしまうから徹底的に戦わない」という集団選択説でも説明できるという主張に対して、それを積極的に否定しているというわけではありません。と なると、どちらを採用するかは個人の好みの問題になってしまいます。これに対して遺伝子選択説派は、次のような例について集団選択説ではどう説明するのかと迫 ります。
1頭のオスと複数のメスでハーレムを構成する種があります。1962年、京都大学霊長類研究所の杉山幸丸は、そうしたハーレムを構成する霊長類ハヌマーン
ラ
ングールを観察していたとき、あるハーレムが他のオスに乗っ取られたとき、その乗っ取ったオスがそこにいた子供たちをすべて殺してしまうという光景を目の当た
りにし、その状況を学会で報告しました。しかし、ローレンツの「種にとっての善」から同種どうしは殺し合わないというのが当時の定説でしたので、それはそのオ
ス特異な異常行動であり、一般性はないと結論づけられてしまいました。しかしその後、ハーレムを構成する種ではよく見られる現象であることが確認されます。
ハーレムでなくても、繁殖期には発情したオスのヒグマが子連れのメスを付け回し、メスが連れている他のオスの子を殺し、メスと繁殖しようとすることが知られて
います。こうした同種の子殺し行動は集団選択説ではどのように説明できるでしょうか。
遺伝子選択説では次のように説明することができます。いま、ハーレムを乗っ取ったとき、前のオスの子を殺すという行動の遺伝子と、殺さないという行動の遺
伝
子があったとします。このとき、殺さないとする遺伝子は、それをもったオスが前のオスの子の育児に参加し、自分の子供を作らないとすれば、子孫には伝わりませ
ん。一方、殺すという遺伝子は、代わりに自分の子供をもうけることになり、子孫に伝えることになります。その行動パターンを引き起こす遺伝子を数多く子孫に伝
えることができるかどうかがこの議論のポイントであって、「種にとっての善」とか、倫理的な論理で説明しようとすると破綻してしまうことを端的に示している、
というのが遺伝子選択説の主張です。
前回の講義で述べた、親が自らの生存率を下げるかもしれない犠牲的な行動で子を守ることも、「種にとっての善」とする集団選択説でも説明できますし、遺伝
子
選択説でも説明できます。遺伝子選択説では、自らの生存率を下げても子を守れという遺伝子は、それによってその遺伝子を引き継いでいる子の生存率を高めること
で、そうした行動を指示する遺伝子自身の生存率を上げることになる、と説明されます。
結局、集団選択説でも遺伝子選択説でも納得できる現象もありますが、すべての現象をまったく同じ論理で説明できるという点で、遺伝子選択説はより合理的で
あ り、優れていると考えられています。
このモデルでは、自然選択における環境への適応ということも考えさせてくれます。通常、環境というと自然環境が頭に浮かびます。また、他種と捕食者‐被捕
食
者の関係にあったり、共生関係にあったりする場合には、他種の生物も環境となります。しかしここに掲げた例では、同種の同一集団に属する他の個体も進化を考え
る上での環境となり得ることを示しています。彼らは、自身と同じ環境を好み、同じ食の好みをもち、配偶者をめぐって争わなければならないライバルであり、生存
率に重大な影響を与える環境なのです。さらに厄介なことは、自分自身も彼らにとっての環境であるため、彼らに変異が起こればそれは自分の子孫に影響を与え、自
分の子孫がそれに呼応して変異を起こすとそれが今度は彼らの子孫に影響を与えるというフィードバック・ループを形成するため、より解析を難しくしています。上
記のモデルは非常に簡略化したものですが、より実際に近いモデルを構築しようとすると、種の特異性なども加味しなくてはならず、一般論を構築するのはきわめて
難しくなることは想像できるかと思います。進化の論理が一筋縄では理解できない理由の一端がこうした個と集団の関係にあることはぜひ心に留めておいていただけ
ればと思います。
自然選択に関わる環境として、同種同群の他の個体も考える必要があることがわかりました。その意味では、異性の個体は特別の意味をもちます。有性の種で
は、
配偶者を得ることは遺伝子を次の世代に伝えるために必須だからです。そのため、性選択とよばれる機構が自然選択のなかで働きます。これまで述べてきた選択機構
とはかなり異なりますので、ここで紹介したいと思います。
オスとメスでは姿が全く違う生物がたくさんいます。クジャクの飾り羽や鳥のオスの派手な色、シカのオスの立派な角などが例として挙げられます。オスのク ジャ クの長い飾り羽は邪魔になりそうで、とても生存に適しているとは思えませんし、オスの派手な色は、メスが地味で保護色を呈しているのを見ると、天敵に対しても 目立ち、生存率を下げる方向に作用しそうです。オスのシカの立派な角も、他のオスとの闘いに優位に立つためとはいえ、ちょっと度を超しているようにも思えま す。そもそも、なぜオスだけがこうした極端な方向に進化が進んだのでしょうか。

性選択とは、配偶者を得る上で有利な形質は、たとえそれが生存には不利であったとしても進化する、そんな機構を生み出すものとして考えられたもの です。そして、派手な姿や装飾を持っているのはオスばかりで、メスは持っていない理由はトリバースの理論によって次のように説明されています。
オスはメスに比べ繁殖に対して大きな投資をしないので、その繁殖成功度は交尾をしたメスの数に大きく依存します。それに対してメスは、子供を育て上げるの に 時間をかけますし、大きな投資をします。したがって、たくさんのオスと交尾をすることよりも、繁殖に必要な時間や資源をみつけることが重要となります。した がって、交尾相手を得られるかどうかはメスにとってよりも、オスにとってより重要な問題となります。多数のライバルオスのなかからいかにメスに選んでもらう か、その答えが、派手な羽や大きな角などだというのです。一方、メスにとっては、どのようなオスを選ぶかがきわめて重要であり、そのため慎重になります。戦略 の一つは人気のあるオスを選ぶことです。そうしたオスの遺伝子をもつ子を産むことは、子がオスのとき、メスに選択してもらう確率を高め、それが子孫を数多く残 すことに大きく寄与するからです。
それではもてるオスの要素(形質)とは何でしょうか。これにはいろいろな説があります。
まずは、メスがオスを選ぶときのポイントはオスが適応度が高いことを示す形質をもつか否かであるとする説です。たとえば、鳥の羽根は寄生虫がついたり、炎 症 が起きていたり、病気をすればすぐに汚くなってしまうため、美しく保たれた羽根はオスが健康であることの証明書となるというものです。すなわち、そうしたオス を選ぶことは、その遺伝子を受け継いだ病気になりにくい、健康な子供を産むことにつながり、メスにとって有利であるというものです。また、オスにとっては、派 手な色、大きな羽根はそれをより強調し、メスによりアピールすることになります。
しかし、そうはいっても、クジャクの長い飾り羽は生存には適さないように思われます。そこでイスラエルの進化学者ザハヴィは、ハンディキャップ仮説とよば れる ちょっと風変わりな仮説を提唱しています。長くて重い羽根や極端に大きな角のように、邪魔なものをつけていると天敵からは逃げにくいし、生き延びるのが難しい だろう。だとするならば、逆に、邪魔な尾羽や角を付けていても生き延びているオスは、そのハンディをものともしないほどに元気であるに違いない。すなわち、ハ ンディと思えるほどの長い尾羽やシカの大きな角は、生存能力の高さや健康であることをメスにアピールする証明書となるというのです。
こうした健康であることや生存能力の高さをメスにアピールするためにオスはさまざまなある種特異な形質をもつという考え方とは異なり、そもそもメスは適応 度の高さにはまったく興味はないという別の視点からの説明もあります。
まず、オスの形質だけでなく、メスのオスに対する好みも遺伝子に支配された生得的なものであると考えます。たとえばクジャクで、オスの飾り羽の長さに対し て、生まれつき短い飾り羽が好みのメスをいれば、長い飾り羽が好みのメスもいるとします。そして、飾り羽が長い方が、上で述べたような意味でより健康で生存率 が高いと、長い飾り羽をもつオスが増えると同時に、長い飾り羽を好む遺伝子をもったメスも増えていきます。こうしてひとたび長い飾り羽をもったオスを好む遺伝 子がメスの間に広まると、より長い飾り羽をもったオスが選ばれるようになり、オスの飾り羽はどんどん長くなっていくというわけです。これをランナウェイ仮説と いいます。当初は飾り羽の長短が適応度という観点から意味のある形質の違いであったかもしれませんが、いったんこの暴走が始まると、そうした意義は失われ、よ り装飾的で非実用的な方向へと進化を推し進めてしまうというのです。さすがに生存に不利なところまで行きつくとこの暴走は止まりますが、メスの選択基準が良質 なオスを選ぶためという論理はもはや通用しない、こんな自然選択の仕組みもあるというのです。
進化では自然選択が働きます。もしその選択機構が強く働いた場合、少数の種へと収束していかないのでしょうか。そうならない理由の一つは、変異の発生が不 可 避であること、そしてその中には自然選択には中立な変異が多数あることです。しかし、この多様性の創出と選択機構のバランスはどのように取られているのでしょ うか。変異の発生を徹底的に抑え込み、最適者とよべるような少数の種で構成する生態系が構築されるのが本来理想であるが、変異の発生から逃れられないために、 仕方なく多様性を生じているということなのでしょうか。それとも、多様性を保持することにこそ重要な意味があり、進化は積極的に多様性を受け入れてきたと解釈 するべきなのでしょうか。多くの生態学者は後者だと信じています。しかし、それを理論的に証明するのはなかなか難しいようです。
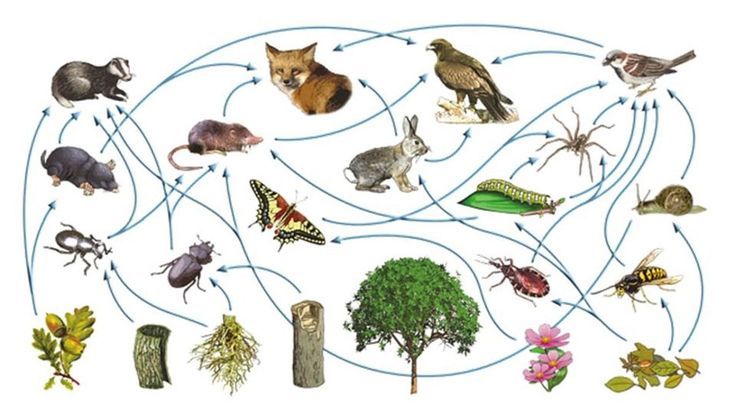
生物多様性を守ろうと叫ばれています。でもなぜそうしなければならないのでしょうか。種の絶滅が起こるということが環境破壊の指標となるので、実は環境問 題 に主眼があって、それを別の視点から注意喚起しているにすぎないとか、まだわれわれが解明していない生物資源が失われてしまうことへの危惧であるとか、消えゆ く生物種への郷愁であるとか、いろいろな解釈がありますが、本質はどこにあるのでしょうか。環境省のホームページには次のように書かれています。「生物多様性 とは、生きものたちの豊かな個性とつながりのこと。地球上の生きものは40億年という長い歴史の中で、さまざまな環境に適応して進化し、3,000万種ともい われる多様な生きものが生まれました。これらの生命は一つひとつに個性があり、全て直接に、間接的に支えあって生きています」。すなわち、地球生態系のなかで 多様な生物が行う多様な相互作用(たとえば食物連鎖;図32.2)あるいはそれによって作られる種間ネットワークがもたらす人類への恩恵こそが生物多様性の本 質だと 言いたいのだと思うのですが、生物多様性に限らず、こうした多様な要素が複雑に絡み合い、ネットワークを構成している系の意義や仕組みについて、自然科学はま だ十分理解できていません。集団は個の集まりであって、したがって個々の要素の性質が集団の性質に寄与することは明らかですが、一方、個々の要素はその集団が もつ性質に影響を受けて変化し、それがまた集団の性質に影響を与えるというフィードバック機構があるため、その解析が非常に難しいのです。そのため現状では、 どうしても漠然と、多様性は大切だよ、という以上のメッセージが出せないでいるのです。
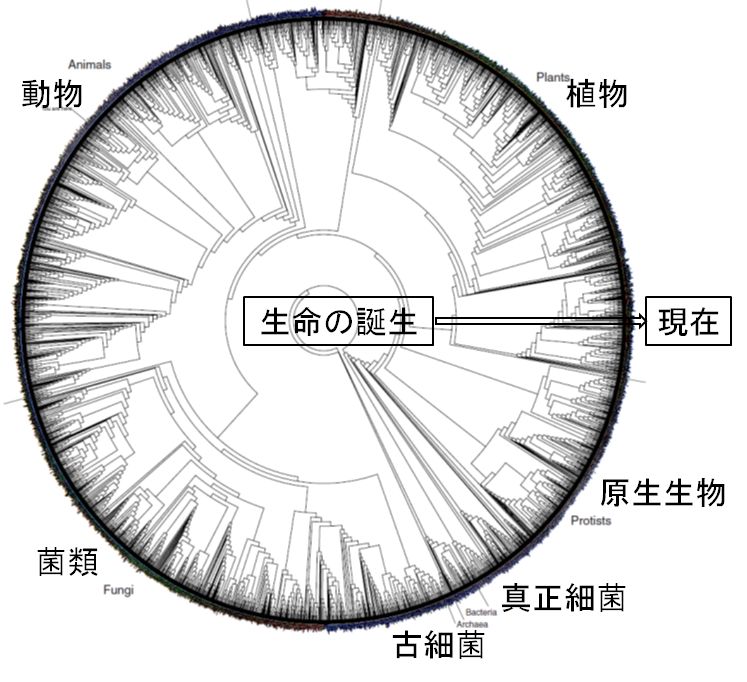
進化は自然選択によって環境に適したものが選ばれることで起こったと語られます。そして単純な生物から複雑・高等な種が誕生した過程が進化だと考えられて き ました。系統樹も、その過程に沿って描かれ、ヒトがその最上段に君臨しているかのように描かれます。しかし、系統樹の下の方に描かれた種の子孫たちも、現在こ の地球上で生きています。彼らをどう捉えたらいいのでしょうか。ある研究者がこんな提案をしました。多様性を生み出してきた過程こそ進化であることを強調する ために系統樹を円で描いたらいいと(図32.3)。これに対して、皆さんはどのような感想をもたれるでしょうか。
進化についてはまだまだ語るべきことがたくさんあります。しかし長くなりすぎたようです。この辺りで終わりとしたいと思います。みなさんは進化をどのように 捉えているでしょうか。ダイバーシティ(多様性)は社会科学にとっても重要だと認識されています。しかしなぜ重要かとなるとそれこそ多様な議論が展開されてい ます。生物の進化と社会の進化とははっきりと分けて論じなければいけませんし、進化と進歩は同じではないことも認識する必要があるでしょう。したがって、ここ で述べたことをそのまま社会科学の問題に適用することはできません。しかし、集団内に多様性を生み出し、構成要員の相互作用によって創発的に形成される何らか の組織、秩序、構造があることを考えてみることは、社会科学を学ぶ皆さんに対しても多くの示唆を与えてくれるものと思います。この講義を契機に、そうした問題 について考えてみるのもいいのではないでしょうか。
ここまでお付き合いいただきましてありがとうございました。
![]()