図31.1ラマルクの進化論
進化というと「下等な生物から高等な生物がどのように生まれたか」が主要な課題と考えられがちですが、この地球上に「なぜこれだけ多様な種が存在するの
か」も重要な視点です。これらの問いは、自然科学だけでなく、社会科学にとっても興味のあるテーマだと思います。もっと一般的な問い方をすれば、最初の問い
は、創造主やデザイナーのような外的にシステムを制御するものの存在なくして、システムが自発的に組織化されるような現象(自己組織化)の仕組みとは何かを問
う
ことですし、2番目の問いは、システムが多様性を内包することの意義を問うことになるからです。今回および次回の講義では、こうした進化を論じるときのさまざ
まな視点に関する話題をいくつかを取り上げてみることにしました。その多くは、まだ確立した理論が得られていませんので、みなさんも自由な発想で、社会科学と
の関連性なども思い浮かべながら、いろいろと考えを巡らせてみていただければと思います。
創造説に基づけば、生物の種は神が個々に創造し、現在に至っていると考えられています。分類学の父と称されるカール・フォン・リンネ(1707 -
1778)は、その著『自然の体系』で生物の分類学を体系化しましたが、種は神が創造したものであり、自然の秩序はその神の意思の現れであると考え,論理的な整合性をもっ
た
分類体系を構築することで、種創造における神の意思を明らかにしようとしたと言われています。冒頭の2つの問いについていえば、種の多様性は創造時に個々の生物種に託した
神の意思の発露であるとし、生物が時間とともに変化し、多様性が生まれてきたという発想はそこにはなかったということになります。
ヨーロッパの大航海時代(15世紀半ばから17世紀半ば)以降、世界各地で新種の動物・植物・鉱物が相次いで発見され、それを分類する手段としての博物学
を
大いに発達させることになりました。しかし、想像以上の種の多様性が明らかになってくると、疑問も湧いてきました。たとえば、すでに知られていた種と種の中間種
が次々と見つかると、種と種の間がより連続的につながることが明らかになり、これらすべての種を何らかの意図をもって最初から創造したのだろうかと。17世紀
末には恐竜の化石も発見されました。大洪水で絶滅したと考えられましたが、過去に生存し、現存しない生物種がいることが明らかになると、彼らは創造主の失敗作
だったのかという疑念も湧きました。でもそれは、全知全能の創造主への冒涜ともいえる解釈です。さらには品種改良も行われるようになり、人為的に生物を変化さ
せ
ることが可能であることもわかってきました。こうして、18世紀の終わりから19世紀の初め頃には、生物種は、創造主による創造以来変わらないのではなく、基本
的な種は創造主によるものだとしても、その後変化したのではないか、あるいは生物種は変化できるという考えが広まるようになりました。そうなると、次の問
いは、どのようなメカニズムで種が変化するのかであり、その理論を構築することが課題となりました。
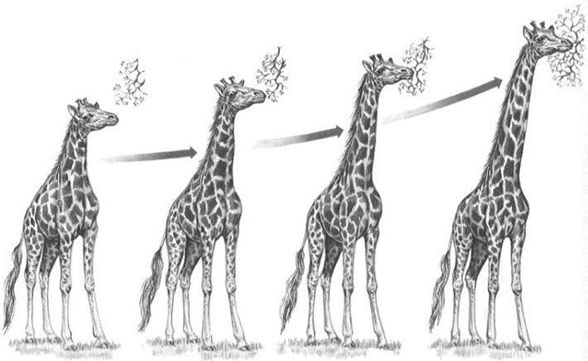
最初に受け入れられたのはフランスのラマルク(1744-1829)が提唱した進化論でした(『動物哲学』1809)。
キリンの首がなぜ長いのかを説明する彼の説明が有名です。元々は短かったキリンの首ですが、下草が少ない環境にいたため、高い枝にある木の葉を食べようといつ
も首を伸ばして努力していました。そして、努力の甲斐あって、成長とともに次第に首が長くなりました。そのようなキリンが子供を生むと、その努力の結果獲得し
た形
質が子供にも伝わり、親が生まれたときよりも少しだけ首が長くなって生まれてきました。これが世代を経て何年にもわたって繰り返された結果、キリンの首は現在
の長さへと変化した、というのです。
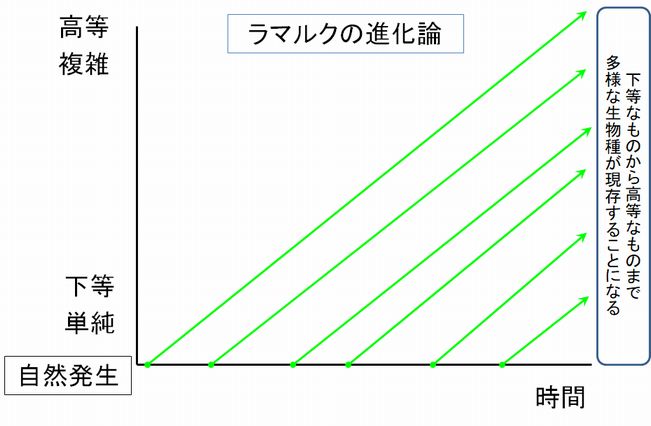
ラマルクは、生物は発展し、環境に適応しようとする目的をその内に有しており、それを実現するために日々努力することによって、単純なものから高等なものへ
と 変
化していくと考えたのです。したがって、生物は、世代を経るごとに進化するものであり、現在複雑・高等な構造をもつ生物はより古い時代に自然発生したからであ
り、現在下等・単純な生物はごく最近自然発生したばかりで、これから高等・複雑なものに変化していくと捉えていました。なお、最も下等・単純な生物は自然発生
するというのが当時の通説でした(パスツールにより自然発生説が否定されたのは1861年のことです)。
こうした考え方はもちろん現在では否定されていますが、生物自身の努力による前進的な進化観、そして親の世代の努力が子の世代へと遺伝するという獲得形質の遺
伝という考え方に共感する人は多く、この後説明するダーウィンの進化論が定説となった後も、長い間信奉者が絶えませんでした。
(注) 親の世代に獲得した形質が次の世代に伝わるかは常に議論されてきました。その理由の一つは、生物の進化と社会の進化とが混同して議論されてきたことが一因となって
います。明らかに社会の進化においては、前の世代が努力して獲得した物質的あるいは精神的遺産を次の世代に継承していきますので、生物の進化にもそうした仕組
みは
あるだろう、あったらいいなあ、という希望がラマルク信奉者を生み出したといえます。しかし、遺伝情報がDNAに塩基配列として書かれており、塩基配列が偶発
的な変異を起こすことはあっても、それぞれの個体の努力によって獲得した形質に呼応して塩基配列が書き換えられる証拠はないと分かっている現在、獲得形質の遺
伝は完全に否定されています。
ところが、最近新たな事実がわかってきています。親の世代に、塩基配列の変化をともわない表現型 の変化が起こることがあり、それが子供に遺伝することがあることがわかってきたの です。親から受け継いだDNAは、その塩基配列を保ったまま、メチル化などの化学的修飾を後天的に受け、それが遺伝子の発現の制御に関与していることがわかっ ています。これはDNAのエピジェネティックな(後天的なという意味)制御とよばれますが、これには各個体の後天的な経験が関与することもあり、クローンとい えども、遺伝子の発現の仕方が異なることがあります。こうしたエピジェネティックな変化は、次の世代にDNAが受け渡されるときに基本的にはリセットされるの で す が、中には受け継がれるものがあることが報告されており、ある種の獲得形質の遺伝ということになり、話題となっているのです。しかし、ある目的に向かって努力 して獲得した形質が遺伝するというラマルクの趣意とは本質的に異なるものです。
 現在の進化論の基礎を築いたのは、皆さんもよくご存じのイギリスの博物学者ダーウィン(1809-1882)です。1859年に出版した著『種の起源』に
より一躍世に知られるようになりました。
現在の進化論の基礎を築いたのは、皆さんもよくご存じのイギリスの博物学者ダーウィン(1809-1882)です。1859年に出版した著『種の起源』に
より一躍世に知られるようになりました。
ダーウィンの時代、まだ遺伝子という発想はありませんでしたし、遺伝の仕組みについてもほとんど分かっていませんでしたので、現代の知識を基盤にした用語
を
用いて彼の理論を説明したいと思います。(遺伝の法則として知られるメンデルの法則は1865年に報告されていますが、1900年に再発見されるまではほとん
ど注目されず、ダーウィンも知らなかったようです。)
進化のダーウィン・モデル
事実1 ネズミ算的に増えるという言葉があるように、生物の個体数は指数関数的に
増加する潜在的な可能性をもっています。
事実2 しかし、多くの種において実際に観測される個体数は、変動はあるものの、
長期的に見れば安定平衡状態にあり、指数関数的増加は観測されていません。
事実3 一方、生物が生存するために必要な食料や住む場所などの資源は有限であり、
指数関数的に増加する個体をすべて養い、収容する能力はありません。
↓
推論1 その結果、種内部において、生存をめぐって個体間の生存競争が起こっていると考えられます。
事実4 同じ種といえども、それぞれの個体には個性(多型)があります。
事実5 そうした個体の個性の中には遺伝するものがあります。
↓
推論2 個体間の違いの中には生き残る確率に差を生み、次世代に残す子の数で差を生み出すものもある
と考えられます。すなわち、遺伝子の違いによる環境への適応度の差が、生存率の差、次の世代を
生む年齢まで育つ子の数の差を生み出すと考えられます。
↓
推論3 たとえその差が小さくても、多数の世代が経過すると、環境への適応度が高い遺伝子をもつ個体
が集団内で多数派を占めるようになりま す。
この中で、遺伝子の環境への適応度の差により自身の生存確率や次世代に残せる子の数に差が生じ、世代を経て集団内に広がるものもあれば、やがて消滅してしま
うものもあることを「自然選択」という言葉で表現しました。
ダーウィンはまた、種が“分岐”して多様な種が生じる、という説を初めて提唱しています。ある生物種の集団が分裂し、棲息する環境が異なったりしたために
お
互いに生殖的に隔離され、それぞれが独立に環境に適応するように変化すると、やがて両者は異なる種とみなされるほどに変貌してしまうというものです。これによ
り、ダーウィンは、創造説やラマルクの進化論のような目的論的な説明を否定し、生物種の多様性が生まれる機構の機械論的な説明を初めて提唱したという点でも特
筆 されます。
いまでも生物学では、目的論的な言説が満ち溢れています。キリンの首は高いところにある木の葉を食べる「ために」長い。カツオは高速で泳ぐ「ために」流線
形 を
している。足は歩く「ために」ある。などなど、生命の合目的的な形質を「~のために~である」と説明することをわれわれは特に違和感を抱くことなく受け入れて
います。しかし考えてみれば、これはその目的を達成するために働く実体について、何かを暗黙に想定している表現になっています。生命に関することになると、信
仰
心がなくとも、神からもらった〇〇という表現を違和感なく使うのも、そうした実体を暗黙に想定していることになります。それに対してダーウィンは、そうした合目的
的にみえる生命現象を偶発的に起こる変異と自然選択というメカニズムで説明しようとしたという点は、しっかりと心に留めておく必要が あります。
ダーウィンの進化モデルは、現在の進化学の根幹をなすものです。しかし、より詳細に検討するとさまざまな課題があることもわかります。進化は実際に観測でき
るわけではありませんので、どうしても不確定性が残るのは仕方のないことでしょう。そのため現代進化学は、化石や現存する生物のゲノムデータを矛盾なく説明で
きる論理の構築を目指すのが一つの方向となっています。そこで、そうした論理構築のなかで重要なポイントとなる視点のいくつかを、ここ
で紹介したいと思います。あ
る程度の意見の一致が見られているものもありますし、議論がまだ続行中のものもあります。進化の論理は社会科学にとっても興味ある話題を含みますので、ここで
提供された話題をぜひ考えてみていただきたいと思っています。
種は変化するといったとき、新たな変異の発生に何らかの法則があるのでしょうか。歴史的には主として次の3つが考えられています。
① 必然性、あるいは定向性。
1960~70年代を代表する日本の進化学者今西錦司の「種は変わるべくして変わる」という言葉に代表されるように、種の変異には、必然性あるいはある方向
を目指した定向性を認めるというものです。
② 獲得形質の遺伝。
ラマルクに代表されるように、それぞれの生き物に内在する適応への強い意思が変化を一定方向へと向かわせるとするものです。定向性という点では、上記①に含
まれます。
③ 変異はランダムであり、変異に先見性はない。
遺伝情報担体であるDNAに起こる変異が種の変異であり、DNAの変異は偶発的出来事である。そこに何ら方向性はなく、将来を見越した先見性もないという
の が現代進化学の立場です。
ここで、①や②のように考えたくなるのにはそれなりの理由があります。一つには、偶発的に起こった変異がすべて受け入れられるわけではありません。それぞ
れ
の種で許容される変異はそのときどきの生体システムや環境に依存し、その意味で、許容される変異はある程度制限されています。さらに進化では、許容された変異
のすべてが化石として残るわけではありませんし、許容されたからといっても集団に固定するかどうかはわかりません。そのため、許容されなかった変異はもちろん
のこと、一時的に存在した変異の多くもわれわれは知る由がありません。したがって、化石などから得られるデータだけから進化の歴史を眺めると、一本道を進行し
て きたように見えるのは致し方ないのかもしれません。
このことは、翻せば、もう一度進化を0からスタートさせたとき、再び現在のヒトが誕生するかと言えば、その確率はほとんど0に等しく、たとえ知能の高い宇
宙人がいたとしても、それがヒトと似たような姿をしている確率もほとんど0に近いということでもあります。
(注) 進化学の中には「前適応」という用語があります。環境条件が変化した際,それに適応するような変異があらかじめ生じていたと
き,その生物は前適応して
いたと表現します。遺伝子重複によりまったく新規な機能をもつ遺伝子が生まれたときなどがこれにあたります。しかし、環境に適応するためにそうした機能をもつ
遺伝子を遺伝子重複によって生み出したというわけではなく、偶然起こった遺伝子重複に、偶発的な変異が起こり、結果的にそのときの環境に適した遺伝子となった
というにすぎません。すなわち、あくまで既存のものの偶発的な「使い回し」であって、その環境変化を予測して作られたものではありません。進化学のなかにはそ
う した学問の歴史を背負った用語もあり、その使い方には注意が必要です。
(注) 新型コロナウイルスの感染力が強まったとき、ウイルスは変異すると感染力を強める方向に進
化する傾向
があるからと解説する人がいますが、これは上記の①や②の論理に相当し、気を付けて聞く必要があります。偶発的にさまざまな変異が起こっているわけですが、感染力を
強めるような変異を起こしたウイルスは、その強さゆえに感染拡大を引き起こしているにすぎず、ウイルスに進化の方向性への意思があるような表現は科学的には不
適切ということになります。
ある個体に変異が起こっても、それで進化が起こったとは言えません。進化はあくまでも集団における現象であり、変異が集団全体に広まることが必要です。こ
れ
を変異が集団に固定したと表現します。ここで集団に変異が広まるとは、感染症のように伝播するわけではなく、世代を通じて変異が受け継がれ、何世代かを経たの
ち、その変異をもった子孫がその変異をもたない子孫よりも、その集団の中で多数派を占めることを意味します。
変異が集団に固定する、あるいは固定しない理由としては次の3つのパターンが考えられます。
① 自然選択に対して中立な変異。
変異が固定するか否かは確率的となります。
② 自然選択に対して保守的な変異。
有害な変異は淘汰されます。結果として現状維持となります。
③ 自然選択に対して革新的な変異。
適応的な変異で、進化の駆動力となり、新たな種の発生などの要因ともなります。
ここで注意しておきたいことは、有利な遺伝子は必ず集団に固定するといった決定論的な議論が長い間されてきましたが、進化の中立説が認められるようになる
と、②や③においても①のような確率論的側面があることが指摘されています。すなわち、有利な変異といえども、固定する確率は中立な変異より高いものの、あく
まで確率的であり、弱有害な変異でも固定する確率はゼロではないことになります。現代進化学では、進化における偶発性がより前面に押し出されています。
また、ある遺伝子に起こった変異が最終的に集団の個体の100%がもつようになるか、あるいは消え去るかといった見方が主流でしたが、これも現在では、い
くつかの変異が多型として共存するのが当たり前であると認識され、そうした複数の変異の頻度の変化として進化を捉えるのが一般的になっています。
さらに、ある時期まで存在した遺伝子が、ある時期に偽遺伝子化して機能しなくなったとき、従来ならば「退化」したと表現し、「進化」の対立語として用いら
れ
ていましたが、それも一つの変異に過ぎず、特別扱いしません。実際、ヒトとチンパンジーの共通祖先がもっていた遺伝子で、チンパンジーでは今でも機能するもの
の、ヒトでは偽遺伝子化したものがありますし、その逆の遺伝子もあります。結局、起こった変異によって機能が(あるいは機能を失うことが)より環境に適するよ
うになったなどの評価はしますが、高等化したとか、進歩したといった価値観をもちこまないというのが現代進化学の立ち位置かと思います。
進化を理解する上で最も重要で、かつ議論の多い点の一つが「自然選択 natural
selection」(自然淘汰とも訳されます)という用語です。以下に3つの視点から、その論点を整理したいと思います。
① 誰が選択するのか
まず「選択」という言葉からは、その行為を行う何らかの実体が想定されます。しかし自然選択において選択する外的実体がいるわけではありません。単に、集
団
の他の構成員より運がよかったか、特定の状況下で有利である属性をもつことで生存率が高かったために生き残ったというにすぎません。それを選択されたと表現し
ているのです。
また、その選択過程は確率的性格をもち、ランダムではないものの、かといって決定論的でもありません。この種をこういう方向に導きたいといった選択におけ
る先見性もなく、そのときどきの条件で選択される確率が決まる極めて場当たり的なものでもあります。
② 何が選択されるのか
次の問いは、自然選択によって何が選択されるかです。これが一番厄介な問題です。進化はある個体に起こった変異から始まります。そしてその変異が集団に固
定
したとき、進化が起こったと考えます。したがって、常に、遺伝子-個体‐群という階層構造を考慮することなしには進化を論じることはできません。これは、社会
科学で、個と集団の関係を議論するのと同じ類の難しさを内包しており、「複雑系の科学」とよばれる学問分野がとりわけ進化をその研究対象の中心に置き、しかも
まだその理解に四苦八苦しているものでもあります。したがって端的にこの質問に答えることはできません。ここでは、何が問題なのかを指摘するに留め、いくつか
の具体的な考察を次回行いたいと思います。
上でも述べましたが、動物の体の構造や機能に関すること、行動パターンに関することなどを説明するとき、31.3節でも述べましたが、われわれはよく「〇
〇
のため」として納得することがあります。こうした表現は目的論的な表現であり、自然選択においてその選択をする実体を想定するのと同じような意味合いをもって
います。進化のダーウィン・モデルで説明したように、偶発的な変異の発生と、適応度の高い形質をもった個体が生き残ってきた結果とする機械論的な説明が
自然科学的には適切な表現ということになります。
しかし、こうした進化に対する「〇 〇
のため」という説明を機械論的な説明に読み替えるのが難しそうな対象もあります。その一つが動物が示す「利他的な行動」 の
解釈です。たとえば、多くの動物は、餌をめぐって、異性をめぐって、縄張りをめぐって種内で激しい争いを起こします。そのとき、多くの場合、敗者が逃げ出すこ
とで決着がつき、勝者が敗者を殺すことはありません。しかし考えてみれば、同種内の個体は、餌、異性、縄張り等をめぐって常にライバル的存在であり、抹殺して
しまうことは勝者となった個体にとって、子孫を増やす絶好の機会となるはずです。にもかかわらず、同種の個体を殺さない行動を説明するためには、そんなことを
した ら種が滅びてしまう、すなわち「種の保存のため」に同種の個体間では勝者は敗者を殺さないとしか説明できないように思われます。
こうした考え方は群選択説(集団選択説、群淘汰説などともよばれます)とよびますが、一方で、そもそも種が存続しているのは個体が種のために尽くすからで
は
なく、その種を構成する個体が存続している結果に過ぎないとして、あくまで個体の存続を中心に置いた論理で自然選択を説明するべきであるとする「個体選択説」
があります。さらには、個体が選択されるというけれども、個体には寿命があり、いつかは死にます。また子孫が残るといいますが、有性の生物では、必ず配偶者の
遺伝子と混合され、選択された優秀な個体そのものがクローンとして永続的に存在するわけでもありません。そう考えると、個体が選択されるということも論理的に
は矛盾するのではないかという議論が生じます。そのため、永続的に存在するものとして遺伝子が浮かび上がり、進化で選択されているのは遺伝子であるとする「遺
伝子選択説」も唱えられています。しかし、遺伝情報担体であるDNAを取り出してもそれを生命とは呼ぶことはできませんし、それぞれの個体はさまざまな遺伝子
から成り立っていますので、遺伝子が選択されるとはどういうことなのかもはっきりしません。さらには、これらを拡張してそれぞれの階層で起こる自然選択がある
とするマルチレベル選択説も提唱されています。
何が選択されるかは進化を理解する上で極めて重要な問題ですが、まだまだ議論が続いています。この点については次回の講義で、もう少し考えてみたいと思い
ま す。
③ 選択の基準は何か
自然選択を考える上でもう一つ問題となるのは、選択されるときの基準です。一般には適応度がその基準とされます。適応度とは、進化学では、ある生物個体が
そ
の生涯で生んだ次世代の子のうち、繁殖年齢まで成長できた子の数として定義されます。それではどのようなとき適応度が高くなるかと言えば、その個体がその生活
する環境に適応し、生存率が高いときと説明されます。しかし、それだけでは説明できない自然選択の基準が設定されていると考えなければならない事象もありま
す。
たとえば、クジャクのオスの飾り羽根はなぜあれほど長いのかという問題です。明らかに生存に有利であるとは言えません。鳥の多くのオスが派手な色をしてい
る
のも、メスが保護色で地味であることを鑑みると、天敵からの回避という点では明らかに生存率を下げる方向に作用しそうです。こうした事例では、性選択という視
点が必要となります。メスに選ばれない限り、どんなに環境に適応していて生存率が高かろうが子孫は残せません。
また、親が自らの命を危険にさらしてでも子を守ろうとする行動は多くの動物で観察されます。これも、その親の生存率だけを考えると説明することができませ
ん。これについては血縁選択(あるいは血縁淘汰)とよばれる自然選択のメカニズムが考えられています。
しかし、こうして事象ごとに選択の基準を設定して理解していくことは仕方のないことなのでしょうか。すべてを一括して説明できる進化の論理はないのでしょ
う か。
また、環境に適したというときの環境とは何かも重要な視点です。通常、自然環境をイメージすると思いますが、それだけなのでしょうか。
一方、自然選択からは中立な変異があることも学びました。それは集団内に多様性を生み出すことになりますが、それは進化の中でどのような意味をもつので
しょうか。これは、ダーウィンの進化モデルに含まれない新たな視点の一つであり、新たな検討が必要です。
とりあえず、自然選択という用語について思いつく問題点を列記しましたが、今回はここまでとします。次回、さらにこの議論を深めていきたいと思いますので、
しばらく自分なりに考えをいろいろと巡らせてみてください。
![]()