
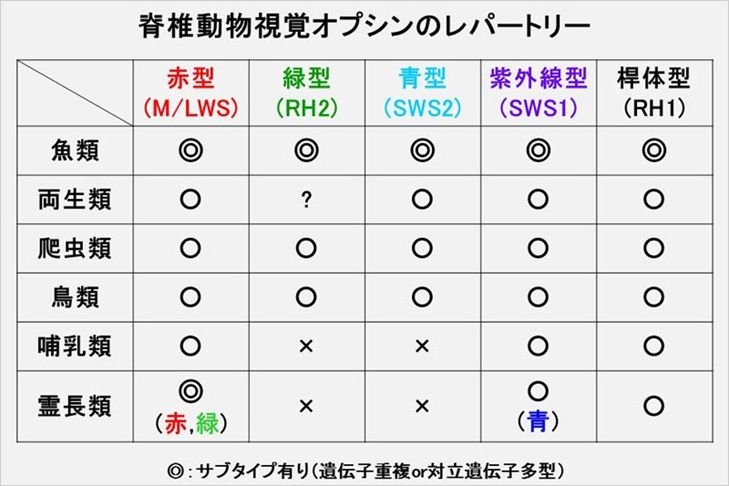
�@�i���͂܂����ׂĂ̐l�ɔF�߂�ꂽ�킯�ł͂���܂���B���ɁA�M���Ă���@���������ے肵�Ă��邩��Ƃ������R�ŐM���Ȃ����������Ǝv���܂����A������ �Ȃ��Ă��A�i�������ۂɌ��邱�Ƃ��ł��܂��A���炩�̈ӎv�����������݂�z�肷�邱�ƂȂ��A�P�Ȃ�����I�ȏo�����Ƃ��Ă��ꂾ�����G�ō����Ȑ����������� �Ă���Ƃ����_�����������Ƃ��m���ł��B
�@�i����ے肷��l�������悭�����o�����Ƃ�������܂��B�u�`���p���W�[���^�C�v���C�^�[�i����Ȃ�p�\�R���� ���傤���j�̌��Ղ��ԁA�����_���ɒ@�������āA���R�V�F�C�N�X�s�A���̍�i��ł��o�����Ƃ��\���낤���v�Ƃ������̂ł��B���炩�ɂ���͐��w�I�ɕs�\ �ł��B�i�������l�ɁA�ł���߂�4��ނ̉����30�����ׂ���q�g�̃Q�m�������R�ł����Ȃ�Ă��Ƃ͐�ɂ��蓾�܂���B���̋^�O�ɑ��āA���㐶���Ȋw�� ������ ���_�ɗ��t����ꂽ�m�ł��铚���������Ă���킯�ł͂���܂���B�������A���Ȃ��Ƃ����̎���̎d��������Ă��邱�Ƃɂ͒��ӂ��K�v�����肻���ł��B���� �̍u�`�͂�����m�M�����Ă���錻�ۂ�������܂��B
�@������DNA��30���̉������Ȃ�܂����A������ڂ������ׂ�ƁA��`�q���܂�DNA�̂���̈悪�d�����đ��݂���Ƃ������ۂ�DNA��̂�����
����
��Ō��邱�Ƃ��ł��܂��B�������`�q�d���Ƃ����܂��B�����̏ꍇ�A�܂����������Ƃ����킯�ł͂Ȃ��A�قȂ��Ă��镔��������̂ł����A���炩�ɁA���X�͂܂�����������
���������̂������N���ŕψق������A�قȂ��Ă����Ɛ���ł��܂��B
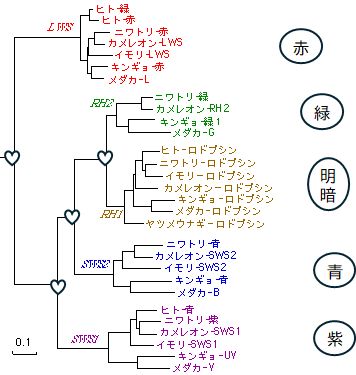
�@��̗�������܂��傤�B���]�`�[���Ƃ���^���p�N���ƃ�-���N�g�A���u�~���Ƃ���^���p�N��������܂��B�ȉ��ɂ��̃A�~�m�_�z�����ׂĕ\�L����
�� �܂����B���������悤�ɁADNA�̉���z�����u����̂Ɗ�{�I�ɓ����Ӗ������������Ă��܂��B
Lysoz MKALIVLGLV-LLSVTVQGKVFERCELARTLKRLGMDGYRGISLANWMCLAKWESGYNTR Album MRFFVPLFLVGILFPAILAKQFTKCELSQLLK--DIDGYGGIALPELICTMFHTSGYDTQ
Lysoz ATNYNAGDRSTDYGIFQINSRYWCNDGKTPGAVNACHLSCSALLQDNIADAVACAKRVVR Album AIVEN--NESTEYGLFQISNKLWCKSSQVPQSRNICDISCDKFLDDDITDDIMCAKKIL-
Lysoz DPQGIRAWVAWRNRCQNR�@ ���]�`�[�� �@lysozyme Album DIKGIDYWLAHKALCTEK ��-���N�g�A���u�~�� ��-lactalbumin
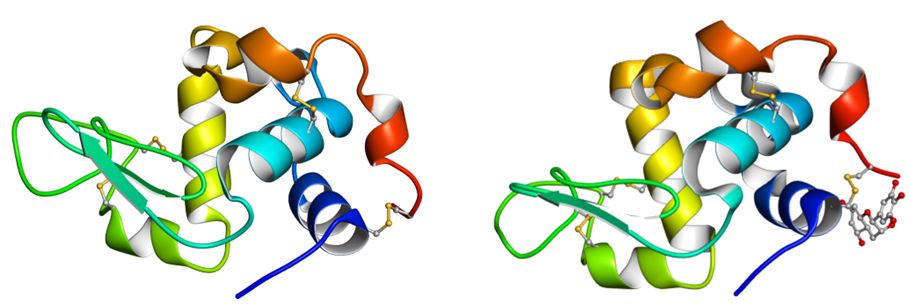
�}30.2�@���]�`�[���ƃ�-���N�g�A���u�~���̃A�~�m�_�z��̔�r
�@�@�@�@�@�@�@�@�@�@
�@�@�@�@�@�@�}30.3�@���]�`�[���i���j�ƃ�-���N�g�A���u�~���i�E�j�̗��̍\���̔�r
�@�e�A���t�@�x�b�g���A�~�m�_�̎�ނ�\���Ă��܂��B���������^���p�N���ŋ��ʂ̃A�~�m�_�A�Ԏ����قȂ�A�~�m�_�ł��B�n�C�t���͑Ή�����A�~�m�_���Ȃ��� �� ���Ӗ����܂��B�����Ő�Ă���Ƃ����킯�ł͂���܂���B���̓�̃^���p�N���̃A�~�m�_�z��̈�v�x�͌����đ傫���Ƃ͌����܂��A����ł��A�܂������W�̂Ȃ� �A�~�m�_�z�����ׂ��ꍇ�A���R�ɂ��ꂾ����v����m���͂���߂ď������A���̈�v�x�͂ƂĂ����R�Ƃ͎v���܂���B���Ƃ��Ƃ͓����^���p�N ���ł��������̂��A�����N���ŕψق��A�قȂ�^���p�N���ɂȂ����Ƃ���̂��ł������I�ȉ��߂ł��傤�B
�@���̂��Ƃ́A���҂̗��̍\�����r���Ă݂�Ɗm�M�ł���ł��傤�i�}30.3�j�B���́A��̃A�~�m�_�z��̔�r�ŁA�Ή�����A�~�m�_���قȂ�ꍇ�ł��A�� �̑��� ���A�������w�I�����̋߂��A�~�m�_�ǂ����ł��邱�Ƃ��قƂ�ǂȂ̂ł��B
�@�Ƃ���ŁA���̂Q�̃^���p�N���̋@�\�����Ă݂܂��傤�B���]�`�[���͐^���ۂ̍זE�ǂ��\�����鑽���ނ�������������y�f�ŁA�q�g�̏ꍇ�A�܁A�@�`�A�����
�ǂɊ܂܂�Ă��܂��B�E�ی��ʂ����邽�߁A���]�`�[���z����搂��Ă��镗�ז������܂��̂ŁA���O�������Ƃ����邩������܂���B�����E�ɍL�����z���Ă�
�� �y�f�ł�����܂��B
�@����A��-���N�g�A���u�~���́A��4�K���N�g�V���g�����X�t�F���[�[ �T
�Ƃ����y�f�Ɖ�̂��`�����ă��N�g�[�X�����y�f�ƂȂ�A���N�g�[�X�i�����j����������@�\�������܂��B���Ȃ킿����̎�v�����̈�ŁA�M���ނɌŗL�̂��̂ł��B
�@�����̒m���𑍍�����ƁA���̂悤�ȃV�i���I��`�����Ƃ��ł��܂��B
�@��R���N�O�A��ނ��������E�̉��҂ł����B���������҂Ƃ����ǂ��q��Ă͗e�Ղł͂���܂���B�����̑c�悽���͔����_�炩�����ɕ���ꂽ�����Y��ł���
��
���B���ɎG�ۂ����荞�ނƒ��̐Ԃ������ł��܂��܂��B�����ŕ�e�����͎E�ە������]�`�[�����܂ފ��̂悤�ȉt�̂ŗ���G�炵�G�ۂ̔ɐB��h���ł��܂�
���B�������̑c��̎q����낤�Ƃ���c�݂̑����ł��B����ȑc�悽���̑̓��Ŏv�������Ȃ��ω����N����n�߂܂����BDNA�ł͒������Ԃ̒��Ŏ��ܕω����N��
��܂��B���鎞�A���R���]�`�[����`�q���R�s�[����A���̌セ�̈ꕔ���ψق����̂ł��B���̌��ʁA��`�q��������^���p�N���̋@�\���ς��܂����B������
�Đ��܂ꂽ�̂���-���N�g�A���u�~���ł��B���R�N�������̏o�����ŗ��̎E�ۂ��ړI��������e�̊��ɊÂ��h�{�����܂܂��悤�ɂȂ����̂ł��B����𗑂��炩
�������Ԃ�����r�߂����ƂŎq��ĂɊv�����N���܂����B�q�����Â����𗊂�Ɉ�悤�ɂȂ�A����̒a���ւƌq�������i���N�����̂ł��B����ɂ���ĕ��
�q�͂�蕪�������������т��A���ꂪ��̈���ւƌq�����Ă������̂ł��B�i���̒i����NHK�X�y�V�����u��������i�Q�@�������ĕ�̈������܂ꂽ�v
(2015) �̉���L �����ꕔ���ς��ėp���܂����j
�@�������������܂��傤�B
�@�O��̍u�`�ł��o�Ă��܂������A�w���O���r���Ƃ����^���p�N���ł��B�Ԍ����ɑ�ʂɊ܂܂�A�x�Ŏ_�f���z�����A�����̍זE�ւƎ_�f���^������@�\��������
��
�܂��B���̃w���O���r���́A����2�{�ƃ���2�{����Ȃ�܂����A�����ƃ����͂��̃A�~�m�_�z���43������v���Ă��܂��B�܂��A�~�I�O���r���Ƃ���^���p
�N��������܂��B�~�I�O���r���͋ؓ��Ȃǂɂ����āA�_�f������@�\�������Ă��܂����A�w���O���r�������ƃA�~�m�_�z��26%��v���Ă��܂��B�����̈�
�v�x�́A���R�ɂ��Ă͂��Ȃ荂���A�������@�\�����̍\�������Ă��邱�Ƃ���A��͂蓯��̑c��^���p�N�������`�q�d���ɂ���Ĕh�����Ă����ƍl�����܂��B
�@���ہA������DNA�ׂĂ݂�ƁA�w���O���r����`�q�Ɋւ��āA�����Ƒ����̈�`�q�d�����ߋ��ɋN�����Ă������Ƃ����肳��܂����B�q�g�ł́A���� �̈�`�q�͑�16�Ԑ��F�̂� �āA�����A�����A���P�A���Q�A���� �Ƃ���d����`�q�Q���A�����͑�11�Ԑ��F�̂ɃÁAG���AA���A�����A�A���Ƃ�� ���d����`�q�Q ���N���X�^�[���`�����Ă��܂��B�܂��A�~�I�O���r���͑�22���F�̂ɂ���܂��B
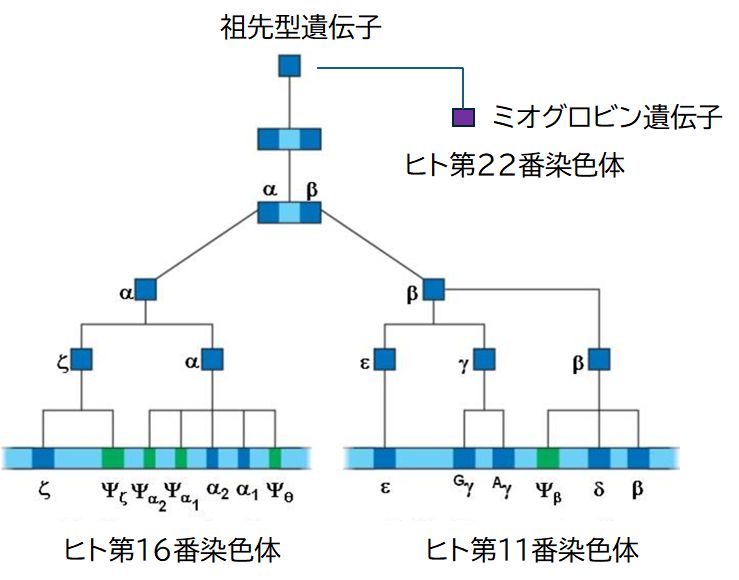
�@�����d����`�q�̉���z��݂͌��ɂ悭���Ă�����̂̂܂����������ł͂Ȃ��A ���ꂼ��قȂ�����������Ă��܂��B�܂��A�����A�����̈�`�q�́A�����A�����A�����A���� �������A�ǂ�����w���O���r�����\������̂ł����A���̔����̎������قȂ� �܂��B�}30.5�Ɏ������悤�ɁA�Ăƃ� ������ɁA���P�A���Q �� G���AA�� �َ͑����ɁA������ ���P�A���Q �� �A�� ���o����ɔ������܂��B�܂��A�@�\�I�ɂ��A�_�f�z���̔\�͂ɍ�������܂��B�َ����ɍ����w���O���r���͏o����ɍ���� �w���O���r���ɔ�r���āA�����_�f�Z�x�ɂ�����_�f�z���\�������̂ł��B�َ��͔x�ċz�����܂���̂ŁA�ٔՂŁA��̂̃w���O���r������𗣂��ꂽ�_�f��َ��̃w���O���r�� �ƌ��������đَ��̑̓��ɉ^��邱�ƂɂȂ�܂��B���̂��߁A�َ��̃w���O���r���̕�����̂̃w���O���r�������_�f�Ƃ̐e�a���������K�v������̂ł��B
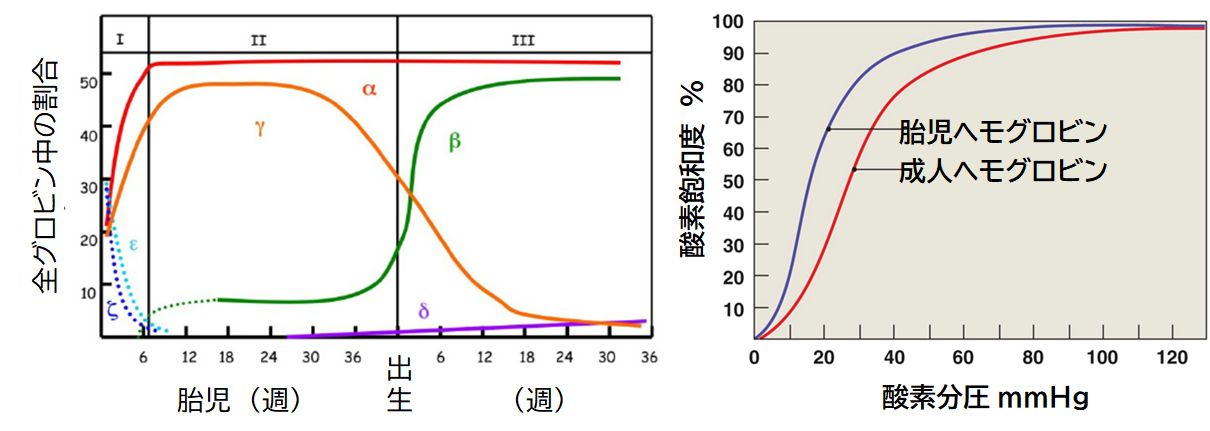
�@�Ȃ��A�����A�����A�����A����
�͋U��`�q�Ƃ��A�@�\���Ă��Ȃ���`�q�ł��B��`�q�d���ł͂���������`�q���a�������Ă��܂��܂��B����ɂ��Ă͎��߂ŐG�ꂽ���Ǝv���܂��B
�@�@
�@��`�q�̏d���́A���F�̂̕s��������g���g�����X�|�]���̓]�ڂȂǂɂ���ċN����A���F�̑S�̂̏d���Ȃǂɂ���Ă��N����܂��B���Ƃ��A���F�̂̕s��
������ �}30.6�̂悤�Ȍo�܂ŋN����A�����̃G���[�Ƃ������ƂɂȂ�܂��B
�@�����̐��B�זE�́A����Ƃ��A���e�����������F�̂ƕ�e�����������F�̂̊ԂŁA�����A���Ȃ킿DNA�̑g�݊������N�����܂��B���̂� ���A���R�Ȃ���A���҂�DNA�̑Ή����镔�������݂��ɐؒf���A�g�ݑւ���K�v������킯�ł����A���ꂪ�~�X���N�����A�}�̂悤�Ɉ���̐��F�̂ɓ�����`�q�� �Q�A��������ɂ͂��̈�`�q�����@����Ƃ������Ƃ��N���蓾�܂��i�s�������j�B�������F�̂͂��ꂼ��ʁX�̐��B�זE�ƂȂ�A�ǂ��炪���邩�͊m���̖� ��ł����A��`�q���d��������������A���ꂪ������ɓ`��邱�ƂƂȂ�܂��B
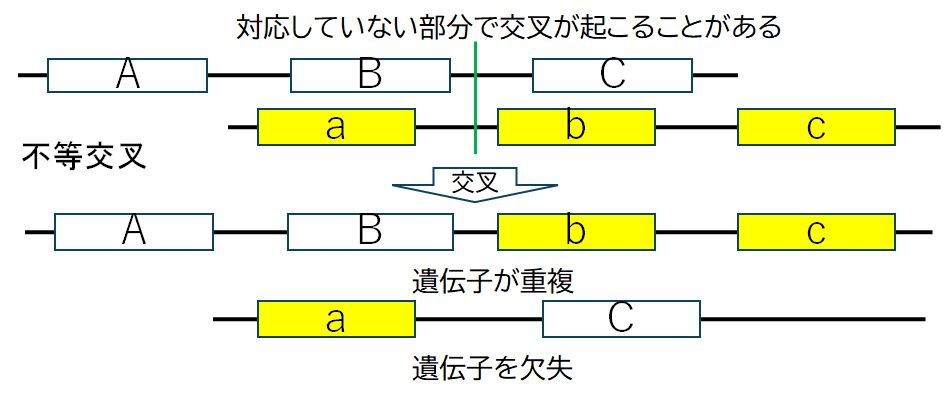
�}30.6�@���e�R���̐��F�̂ƕ�e�R���̐��F�̂͌����Ƃ��錻�ۂɂ���āA�g�ݑւ���
�N�����܂��B�����́A�����݂��ɑΉ�����ӏ��Ő�\����s���܂����A���ꂪ����邱�Ƃ�
����܂��B���̌��ʁA��`�q�̏d������ь������N����܂�
�@��`�q�̏d���ɂ�郁���b�g�͉��ł��傤���H�O��A�f���v���𑽂��ێ悷�閯���ł́A�f���v�������y�f�̈�`�q�̃R�s�[���������Ƃ����b�肪����܂������A
�܂��Ɉ�`�q�d���ɂ����̂ł��B�R�s�[�����������Ƃň�C�ɑ�ʂɂ��̍y�f���������邱�Ƃ��\�ƂȂ�A�f���v�����܂ސH���̏����̉��P�ɂȂ��郁���b�g
������܂��B�������A������������Ƒ傫�ȃ����b�g������܂��B����́A�����S�ɐV�K�@�\����������`�q��n�����邱�Ƃ��\�ƂȂ邱�Ƃł��B
�@����܂ʼn��x���������Ă����悤�ɁADNA�͋����I�ȕψق��瓦��邱�Ƃ͂ł��܂���B�����āA���̕ψقɂ��e���́A�����̏ꍇ�A��`�q�Ƃ��Ă̋@�\��
����
�Ȃǂ̃l�K�e�B�u�Ȃ��̂ŁA�悭�Ă��I�����ɑ��Ē����ȕψقł����āA���K���x�̍�����`�q�ւƕω�����ψفA���邢�͂܂������V���ȋ@�\�������Ƃɂ�
��悤�ȕψق͂���߂Ă܂�ɂ����N����܂���B�������A�܂������V�K�ȋ@�\������`�q�ƂȂ����Ƃ��Ă��A����ɂ���ď]���̈�`�q�̋@�\���������Ƃ��L��
���ǂ���
���킩��܂���B���������뜜���������Ă����̂���`�q�d���Ȃ̂ł��B�R�s�[�������Ƃɂ���āA����̈�`�q�ŏ]���̋@�\��ێ����Ȃ���A��������̈�`
�q�ł��܂��܂Ȏ��݂��\�ƂȂ�̂ł��B
�@�w���O���r���̗�Ō����̂́A��{�I�ȋ@�\�͕ێ����Ȃ���A�������_�f�̋z���\�������قȂ���̂������Ƃɂ���āA�ٔՂ�ʂ��Ă̎_�f�̋�������
�\
�Ƃ��A�ِ��ɂ���Ă��m���Ɏq��Ă��ł���悤�ɂȂ����Ƃ������̂ł����B���̂悤�ɁA�]���̋@�\������`�q��ێ����Ȃ���A�������@�\�̈قȂ�V����
��`�q��lj����Ă������Ƃɂ��A��荂���ȁA��蕡�G�ȃV�X�e��������������̂ݏo���Ă����ƍl�����܂��B����������́A�u30.4
���o�A�k�o�A���o�v�̐߂ł����邱�Ƃ��ł��܂��B
�@�܂��A���]�`�[���ƃ�-���N�g�A���u�~���̗�ł́A�܂������قȂ�@�\����������`�q�ݏo���A���ꂪ�M���ނƂ����V���Ȏ�ݏo���_�@�̈�ƂȂ�
��
�܂��B����������Ƃ��Ă悭�m���Ă�����̂ɁA�N���X�^�����ƔM�V���b�N�^���p�N��������܂��B�M�V���b�N�^���p�N���́A�זE���M�Ȃǂ̃X�g���X�ɎN���ꂽ
�Ƃ��זE��ی삷�邽�߂ɓ����^���p�N���ŁA�����E�ɍL�����z���Ă���^���p�N���ł��B����A�N���X�^�����́A�����̊�ɂ�����튯�A�����̂̍\�������ŁA��
��
��ɂ���Ă��܂��܂Ȏ�ނ̃^���p�N�����g���Ă��܂��B�M�V���b�N�^���p�N���̈�`�q�d���ɂ���Đ������^���p�N���𗘗p�������̂����̈��ŁA��̒a��
�ւƌ��т����ƍl�����Ă��܂��i���҂̐����w�I�@�\�͂܂������قȂ�܂����A�A�~�m�_�z��̗ގ����̍��������`�q�d���ɂ���Ēa���������̂Ɛ��肳���
���܂��j�B�������A�N���X�^�������ォ��a�������킯�ł����A�N���X�^�����̍ő�̖����́A���ꂪ�ÏW�����Ƃ��A�����Y�̖���������悤�ɓ����ł��邱�Ƃ�
��
��A���̏��������^���p�N���Ȃ�A�����炭���ł��悩�����킯�ŁA�M�V���b�N�^���p�N���ɑ����ψق��N���������̂ŏ\���ɖ������ʂ������Ƃ��ł��A���p��
��
���A����Ȋ����ł��傤���B�����ŋ������Ă����������Ƃ́A�i���Ƃ́A�ŏ������̂��Ƃ܂ŏ\���l������Đv���ꂽ���̂ł͂Ȃ��A���R�̕ψق̒��ŁA���ɏ�
������I�ɋN�����Ă����Ƃ������Ƃ��A�����̎����������Ă��邱�Ƃł��B
�@�w���O���r���̗�ł́A��`�q�d�����U��`�q�ݏo�����Ƃ������Ă��܂����B�d����̋����I�ȕψق́A��ɐ�������킯�ł͂���܂���B���s�삪������
�Ĉ�
�`�q�̒��Ɏc����Ă���̂ł��B���Ȃ݂ɁA�����̃Q�m���̒��ɂ́A��`�q���Ə���Ƃ����Ȃ����̋U��`�q������Ɛ��肳��Ă��܂��B���������������ɂ�
�U��`�q�͌���ꂸ�A�������A���G������قǂ��̐����������Ƃ��m���Ă���A��`�q�d�����i���ɂ����ɑ傫�Ȗ������ʂ����Ă��������v���m�点�Ă���܂��B
�@����̍u�`�����̖`���̘b��ɖ߂��Ă݂܂��傤�B���܂���ꂪ����DNA�̉���z��̂��ׂĂ��꒩��[�ɂł����Ƒz�肷��Ƃ���͐�ɕs�\�ł��B��
��
���A���炩�̏������Z��DNA�������N�̊��Ԃ������ċ����I�ɒa�����邱�Ƃ͉\�ł��B���̌�A��`�q�d���ƕψقɂ���āA���X��DNA�̗ʂ𑝂��A���
�ʂ𑝂₵�Ă����ƍl����A�������Ɋ�Â����i�����_�������ĕs�\�ł͂Ȃ��͂��ł��B���Ƃ��A����ꂪ�g������̌n�ł��A�ŏ��͖{���ɂ킸���ȒP���
�X�^�[�g�����͂��ł��B���̌�A�P�ꐔ�������������Ȃ���A�܂�����ɂƂ��Ȃ����@�����X�ɕϖe���Ȃ���A�`��������ʂ𑝂₵�A���݂ւƎ���A���܂�
��
�����ϗe���Ă��܂��B�����悤�Ȃ��Ƃ������̐i���ɂ��N�������Ƒz�肷�邱�Ƃ͌����čr�����m�Ȃ��Ƃł͂Ȃ����Ƃ��������Ă���������ł��傤���B������
��A�����ؖ��͂���Ă��܂��A�����̐������DNA�̗l���������I�ɐ���������̂ł��邱�Ƃ͊m���ł��B
�@1960�N��̈�`�q�d�����������̃p�C�I�j�A�I���݂ł����슣�́A��`�q�d���ɂ��i�����A�u�ŏ��̈�n���A���̌�̕S����v�ƕ\�����Ă��܂��B
�@�Ō�ɁA��`�q�d���̗�Ƃ��āA���o�A�k�o�A���o�Ɋւ���`�q�̘b������������b���������Ǝv���܂��B
(1) ���o�Ɋւ���`�q
�@�q�g�͂R��ނ̐��̎������i�ԐF�A�ΐF�A�F���I�v�V���Ƃ���^���p�N���j�ɂ��R�F�^�F�o�ŐF��F�����Ă��܂��B�e���r��p�\�R���̉�� ���A �ԁA�A��3���F�̑g�ݍ��킹�ő��ʂȐF��\�����Ă���̂͂��̂��߂ł����A�Ƃ�����ƁA�F��3���F�ŕ\�����邱�Ƃ������w�I�ȗv�����Ǝv���Ă��܂������� ���B�������A�����́A����̈�̔g�����������d���g�ł���A���͈̔͂Ŕg���͘A���I�ɕω����Ă��܂��̂ŁA3���F�ŐF��\�����邱�Ƃɉ��畨���w�I�ȕK�R�� �͂���܂���B����ɁA�}30.7�Ŏ������悤�ɁA�q�g���܂ލ����쒷�ނ�3�F�^�F�o�ł����A���̑��̚M���ނ�2�F�^�F�o�A�����̒��ށA��ށA���ނ͂S�F�^ �F�o�A�Ƃ����悤�ɈقȂ��Ă��܂��B�i���I�ɂ݂�A����ꂪ3�F�^�F�o�ł���̂͒P�Ȃ���R�ɉ߂��܂���B
�}30.7�@�Ғœ����̐F�o�B�������A��ԉE�̗�̞��̌^�́A���Â݂̂ɔ������鎋�����ŁA�F�o�ɂ�
�܂܂�܂���B�Z�͂��̃^�C�v���P��ށC���͂Q��� �ȏ�
�����Ă��邱�Ƃ�\���C�~�͎����Ă��Ȃ����Ƃ�\���Ă��܂��B���ނɂ͂T�^�C�v���ׂĂɂ��Ĉ�`�q�d���ɂ��T�u�^�C�v������Ă��܂��B
�@�}30.8�Ɍ���悤�ɁA4���N�O�A�Ғœ�����4�F�^�F�o�������Ă��܂����B���ɔ�������^���p�N������`�q�d�����J��Ԃ��A�ψق��邱�Ƃŋ������������
�̔g��
���������ω����A�������S���Ă����ƍl�����܂��B�������������h���A�M���ނ͖�̐��E�Ő�����悤�ɒǂ������ƁA�F�o���ꕔ�����A2�F�^�ւƕω�����
���B�������A���悻3500���N�O�A�����쒷�ނɂ����āA�Ԍ^�������̈�`�q�ɂӂ����ш�`�q�d�����N����A���̌�ψق��N����A������Ό^�ւƕω����Č���
�Ɏ����Ă��܂��B���Ȃǂ̎��������g���S�̂��ϓ��ɃJ�o�[
���Ă���̂ɑ��āA�q�g�̐Ԍ^�ƗΌ^�̑Ή�����g���͈͂͂��Ȃ�d�Ȃ��Ă��܂��B�݂��ɂ܂����Ă���̎��Ԃ��Z���A�����������Ƃ������Ƃ�
���B���ہA�������̃A�~�m�_�z���364��15��������Ă��܂���B
(2) �k�o�A���o�Ɋւ���`�q
�@�������l�Ԃ̏ꍇ�A���퐶���ōł��S���������o�͎��o�ł����A��Ō����悤�ɁA�O�E�̌������m�������e�̈�`�q�͂�����������ނ�������܂���B��
���A
���o�́A�Ö��A�ꖡ�A�|���A�����A�_����5��ނ���A���ɋꖡ�������Ƃ�ꖡ��e�̈�`�q�͐��\����܂��B����A�k�o�́A�Ғœ��������Ă݂�ƁA���S�����
���S���̎�e�̈�`�q�𑵂��Ă��܂��B����̓Q�m����������S��`�q�̐��p�[�Z���g�ɂ��Ȃ锜��Ȑ��ł��B���o�A�k�o�������ɏd�v�Ȋ��o�ł��邩�����
���܂��B
�@���o��e�̂ƚk�o��e�́i��������^���p�N���ł��j�́A�����Ǝ_���������A�H�a�Ɋ܂܂�鉻�w������V���鉻�w�����ƌ��ƌ����̊W�Ō������邱
��
�Œm�o����_�ŋ��ʂł���A�����̎�ނ𑝂₷���ƂŁA���܂��܂Ȗ���ɂ����ʂ���\�͂����߁A���������グ�Ă����ƍl�����܂��B�������������̑��l��
�́A��`�q�d���ƕψقɂ��B������܂����B
�@�}30.9�́A���܂��܂Ȏ�ɂ��Ěk�o��e�̈�`�q�̐��ƁA���̂����ǂ̂��炢���U��`�q���i��`�q���ψق��A�@�\�������� �Ɓj
���Ă��邩�������Ă���
���B�}�E�X��C�k�ɂƂ��Ěk�o����̏�����ɏd�v���͊F��������������Ǝv���܂����A�����͂��łɚk�o��`�q�̔����ȏオ�U��`�q�����Ă���A�k�o��
��̏��̏d�v�������Ȃ茸���Ă��܂��B�k�o��`�q�̋U��`�q���͂R�F�^�F�o���l���������Ƃ���}���ɐi��ł��܂��B���o���瓾�����k�o���瓾����
�����͂邩�ɏ���悤�ɂȂ������߂ƍl�����Ă��܂��B
�@�Ȃ��A�w���O���r���̋U��`�q�ł́A�@�\������`�q�ɂȂ葹�˂��ƕ\�����܂������A�k�o��`�q�̏ꍇ�́A���Ƃ��Ƃ͋@�\���Ă�������ǂ��A�ψق��N���� �ċ@�\���Ȃ��Ȃ��Ă��x�Ⴊ�Ȃ��������߁A���ɓ�������邱�Ƃ��Ȃ��A�U��`�q�������܂c�����ƍl�����܂��B
�}30.9�@�k�o��e�̈�`�q�̐��ƋU��`�q�̊���
�@�i���̒��œ��M���ׂ��o�����ɃJ���u���A�唚���Ƃ�����̂�����܂��B��10���N�O�ɑ��זE�������a�����܂������A���悻5��4200���N�O����5�� 3000���N�O�̊Ԃɓ˔@�Ƃ��āA�����E�̍ő�̕��ޒP�ʂł���u��v�ɑ������铮�������̑c�悪��Ăɏo�������̂ł��B���̔����I�ȑ��l���̌����ɂ��Ă� �܂������ȓ����͂���܂��A�_�f�Z�x�����݂�20%�ɒB���A�I�]���w���o�������O����������悤�ɂȂ��āA����ւ̐i�o���\�ƂȂ����Ȃǂ̊��̕� �����w�E����Ă��܂��B�V�V�n�ɂ͂܂��V�G�͂��炸�A���l�Ȑ����������\�ł������ƍl�����܂��B
�@����������̑��l���ɂ́A��`�q�d�����傫����^���Ă���킯�ł����A�������鐶���̈�`�q�̉�͂���́A�����I�ɓ����̌`�Ԃ����l�������Ǝv���鎞
���A
���Ȃ킿�J���u���A�����ɂ͈�`�q�̑��l���͂قƂ�nj���ꂸ�A�ނ���10���N�O�`9���N�O�����5���N�O�エ�悻�ꉭ�N�̊Ԃ̂Q�̊��ԁA���Ȃ킿�J���u��
�A�����̐����N�O�ƃJ���u���A������1���N��Ɉ�`�q�d��������ɍs��ꂽ���Ƃ��������Ă��܂����B��`�q���x���̑��l���ƌ`�ԃ��x���̑��l���̎����͖��炩
�ɏd�Ȃ��Ă��Ȃ��̂ł��B
�@���̈�`�q���l���p�^�[������������ł��d�v�ȓ_�́A��`�q�̑��l���̓J���u���A�����̒��ڂ̈������ł͂Ȃ������Ƃ������Ƃł��B�J���u���A�����ƈ�`�q
��
���̎��ԓI����́A�J���u���A�����������N���������q���x���̃��J�j�Y�����l�����ŁA�V������`�q�����Ƃ����n�[�h�̎��_�ł͂Ȃ��A���łɑ��l��������`
�q�������ɗ��p���ăJ���u���A������B���������Ƃ����\�t�g�̎��_���d�v�ł��邱�Ƃ���Ă��܂��B�i�{�c���@�i���̘b�@
https://www.brh.co.jp/research/formerlab/miyata/2006/post_000002.html �Q�Ɓj�@
![]()