タンパク質の形を通して学ぶ「遺伝情報とは」
第29回 ヒトの進化(2)
およそ20万年前のアフリカを発祥とする現生人類は、10万年前から地球全体へと拡散し、熱帯・寒冷地、高地・低地、海辺・山岳地帯などさまざまな地域に
定住し、適応してきました。およそ1万年以内には、現生人類は狩猟・採集から、農耕・牧畜へとその生活様式を変化させ、急速な人口増加、人口密度の増加、そし
てさまざまな感染症の蔓延も招きました。また地域環境への適応や食餌の変化は、その地域独特の遺伝的特性も生み出しています。
近年、各国で数千人、数万人単位で個人のゲノム(DNAの1セットすべて)の塩基配列が解読できるようになり、ゲノム全体にわたって塩基配列を比較するこ
と ができるようになりました。これはゲノムワイド関連解析(Genome Wide Association
Study)とよばれますが、さまざまな形質に関する遺伝子が特定され、しかもある地域で、その遺伝子に起こったある変異をもつ子孫が最近(1万年〜数千年の間に)急速に
増加したと推定されるものもいくつか見つかるようになりました。すなわち、人類の適応進化の様相が遺伝子レベルで徐々に明らかになっています。図1は、
2016年、科学雑誌Scienceに掲載された総説から引用した図ですが、その代表的な例を示しています。
前回、前々回と中立的な進化の話をしましたが、適応的な進化も一方では存在しますので、今回はその話題に目を向けてみましょう。
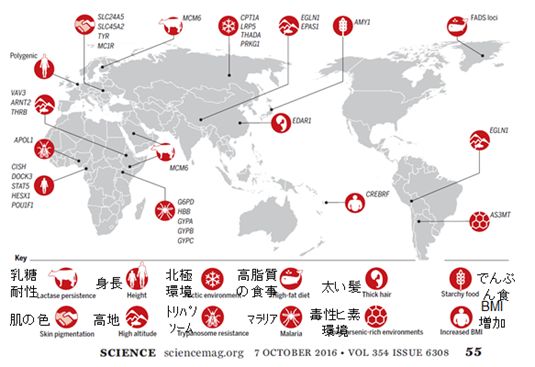
図29.1 さまざまな地域で最近起こった人類の適応進化の例
地図の中の赤丸の絵の横にある文字列は関連する遺伝子名で、その遺伝子に起こったある変異を
もつ人が、ここ数千年から1万年以内に急速に増えてきたと推定されるものが示さ れています。
なお、polygenicとあるのは多数の遺伝子が関与していることを意味します。
図の出典:S. Fan, et al., ”Going global by adapting local: a review of recent
human adaptation”,
Science, 354, 54‐59 (2016)
図29.1には、それぞれの地域で食餌に関連した適応(乳糖耐性、高脂質の食事、でんぷん食)、生活環境に関連した適応(肌の色、北極環境、高地、毒性ヒ
素が多い
環境)、感染症に関連した適応(トリパソーム、マラリア)、身体的な適応(身長、太い髪、BMI増加)が起こった例などが掲げられています。いずれも、その関
連遺伝子が特定されているもので、これらの中からいくつか紹介したいと思います。
なお、すでに取り上げたものには、以下のものがあります。
乳糖耐性 第11回 転写因子 付録 ラクトース不耐症
マラリア耐性 第12回 SNP 12.2(例1)ヘモグロビン
肌の色 第21回 DNAの損傷 付録 肌の色
29.1 高脂質の食事
アラスカ、カナダ、グリーンランドなどの氷雪地帯に住むイヌイットの高脂肪食への適応に関して発表された論文の概要を以下に引用しましょう。
Fumagalli et al. Scice (2015) 349, 1343-1347.
イヌイットは、過酷な気候下に住むため、他のほとんどの民族集団と比べて、タンパク質と脂肪をはるかに多く含む食事を摂っています。今回、イヌイットの遺
伝
子構成の解析により、この集団がどのようにして環境に適応してきたかが明らかになりました。生理学的性質の中で、脂肪代謝、身長、体重、コレステロールの制御
に関わる多数の遺伝子が、おそらく選択圧によって、イヌイット集団を、タンパク質と脂肪を多く含む食事で生きられるようにしたことが判明したのです。
イヌイット集団の遺伝子構成に関する更なる洞察を得るため、ヨーロッパ系の先祖を持つ割合が5%未満のグリーンランドのイヌイット191名のDNAを解析
し、結果を、ヨーロッパ系および漢民族系の人と比較しました。そして、ω6およびω3脂肪酸を不飽和度の高い、長い脂肪に変換することを制御している酵素の遺
伝子
など、多数の興味深い遺伝子を同定しました。さらなる解析により、これらの遺伝子の進化的選択が、イヌイット集団がネイティブアメリカンから分岐する前に開始
されたことも明らかになりました。このとき、彼らの共通の祖先は、ベーリング海峡(ネイティブアメリカンの祖先がアジアから北米に渡るために利用した陸の
「橋」)周辺に住んでいました。
解析で目立った他の遺伝子としては、高脂肪の摂取に伴う酸化ストレスを防御する遺伝子、心筋症に関連する遺伝子、高脂肪食とそれに関連したインスリン抵抗
性による肥満に対抗してエネルギー消費を増やす遺伝子、肥満細胞の分化を制御する遺伝子などがありました。これらの結果は、イヌイットが、蛋白質と脂肪に富む
非常に特別な食事を食べて生き延びるための遺伝的多様性を備えていることを示唆しています。
29.2 でんぷん食への適応
遺伝子のコピー数(同じ遺伝子を複数個もつということ)にも多型があることが発見されました。2007年にNature Geneticsに発表された論文(Perry
et al., Nature Genetics (2007) 39, 1256–1260 )の概要でご紹介します。
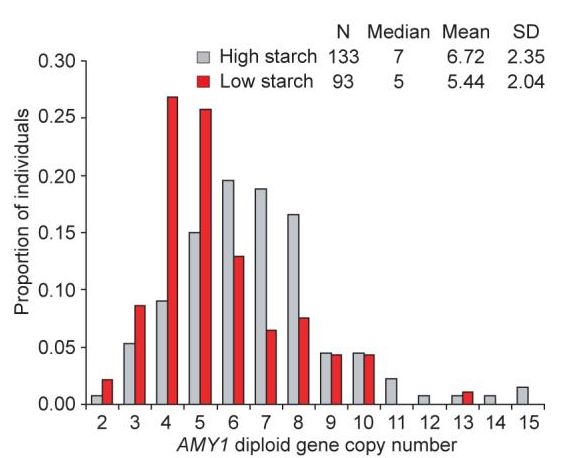
図29 唾液アミラーゼ遺伝子 AMY1 のコピー数の分布
横軸にコピー数、縦軸にその頻度を示している。赤色の棒グラフはデンプンをあまり
摂らない集団、グレーの棒グラフはデンプンを多く摂る集団の頻度分布
デンプンの消費の多さは、乾燥した環境における農耕民族と狩猟採集民族にみられる顕著な特徴です。それに対して、熱帯雨林や北極圏周辺の狩猟採集民族や一部
の
牧畜民族は、はるかに少ない量のデンプンしか消費しません。こうした食行動の相違から、異なる選択圧がデンプンの加水分解を担う酵素であるアミラーゼに働いてい
た可能性が浮上してきました。今回、唾液アミラーゼ遺伝子(AMY1)のコピー数が唾液アミラーゼの発現レベルと正の相関を示すこと、また、デンプンを
多くとる食習慣をもつ集団に属する個体は、平均して、伝統的にデンプンをあまりとらない集団の個体よりも、遺伝子AMY1のコピー数が大きいことを見つけました。
これらの集団のサブセットでその他の遺伝子座と比較したところ、AMY1のコピー数の多型の程度は並外れて大きいことが示唆されました。このように、コピー数
が変動するように進化の過程で選択圧がかかった例は、知られている限り、ヒトのゲノムで最初に発見されたものです。AMY1のコピー数が大きく、それによって
合成されるアミラーゼの量が多くなると、デンプンを含む食物の消化がおそらく改善し、消化管疾患による適応度を低下させる効果を和らげるのではないかと考えら
れます。
29.3 青い目
図29.1には青い目は含まれていませんが、青い目は虹彩のメラニンの量が少なく、長波長の光(赤色の光)を吸収し、短波長の光(青色の光)を反射するた
め青く見え
るので、肌の色とも密接に関連している形質であることがわかっています。肌の色についてはすでに第22回の講義で取り上げましたので、ここでは青い目の遺伝子を
取り上げます。
青い目の遺伝様式は劣性で、複数の遺伝子からなる形質です。2008年、Eibergらは、OCA2遺伝子のプロモーター(遺伝子の前にあって、遺伝子の
発現
を調節しているDNA上の領域)と相互作用していると考えられているHERC2遺伝子のイントロン部分にある変異が、OCA2遺伝子の発現を減少させ、それに
引き続いて起こるメラニンの合成を減少させていることを明らかにしました。そして進化学的な解析から、その最初の変異は
6,000〜10,000年前の黒海周辺のある個人に起こったと結論づけています。
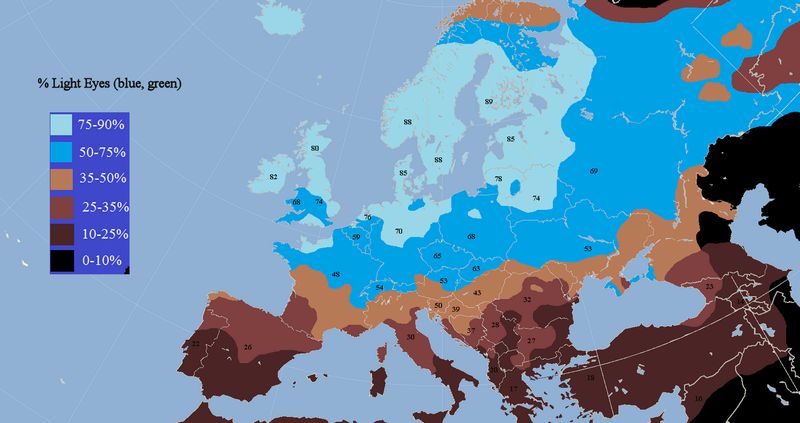
図29.2 青い目の人たちの割合
図の出典:Light Eyes in Europe.png @Wikimedia commons
図29.2は、それぞれの地域での青い目の人たちの割合を示しています。明るい水色の地域が最も割合が高いのですが、黒海周辺で発生した変異は北方へと移
住した 人た
ちでより適応性に影響し、変異をもった子孫たちがそこで繁栄した様子が見て取れます。(前回の講義で、遺伝子の分布を見ると、最も頻度の高いところが発祥の地
で、そこから拡散していった様子がその頻度分布から読み取れると述べましたが、ここではそれが成り立っていません。その遺伝子によって環境に適応することが生
存に大きく関わる場合には、強い進化的な選択が起こり、高頻度の地域が必ずしも発祥の地とは言えない場合があります。ここでは、変異が起きた時代の推定と、ア
フリカを出た人類がヨーロッパへと拡散していった様子などを考慮し、そうした推定がなされています。)
29.4 高地順応
チベットとアンデスの高地に暮らす人々の高地順応に関わる遺伝子について発表された次の論文の概要を引用します。 Lawrence et al.,
Sci. Adv. 10, eadj5661 (2024) ただし、付加情報のある https://newspicks.com/topics
/synbio/posts/249 に掲げられた翻訳をベースにして います。
地理的には離れながら、同じように高地に住むチベットとアンデスの多くの民族(以下、チベット人、アンデス人と呼びます)は、それぞれ独立に、高地での低
酸素条件に耐えられるように順応しています。数多くのゲノム比較研究により、多くの候補遺伝子がこのヒトの高地順応に関係していることが示唆されています。と
りわけ、低酸素に対する細胞応答のマスター転写調節因子である低酸素誘導因子 (HIF) に関係したパスウェイの遺伝子が重要と考えられています。
HIF-2のαサブユニットをコードするEPAS1
は、チベット人およびチベット犬といった高地に適応した動物で多型が見られる遺伝子です。チベット人に見られるEPAS1遺伝子の変異は、転写産物であるEPAS1タンパ
ク質のアミノ酸配列の変異ではなく、遺伝子発現の制御に関わるものであるとされています。これは、チベット人の比較的低いヘマトクリット値(赤血球の全容積が
全血液中に占める割合、つまり血液の「濃さ」を表す指標)に、そしてその他の適応表現型
(過剰な赤血球増加症からの保護など)に間接的または直接的に関与しています。このチベット人の変異は、絶滅した旧人類であるデニソワ人から受け継いだものと考えられてい
ます。
一方、アンデス人には、EPAS1タンパク質のアミノ酸配列の中の1つのアミノ酸の変異が見つかっています(A>G、His 194
Arg)。このアンデス人特有の変異は、アンデス人にはそこそこの頻度で存在しますが、チベット人を含めて、他の人類集団や脊椎動物(シーラカンスを除く)には存在しませ
ん。この変異をもつように人為的にゲノム編集されたヒト細胞は、HIF-2の標的遺伝子の発現が低く、低酸素によって調節される遺伝子発現と同様の変化を示し
ました。また、メタボロミクス分析により、低地集団における遺伝子型と表現型との関連が明らかになりました。それによると、アンデス人に見られるEPAS1タ
ンパク質の1アミノ酸のみを変化させるこの変異は、比較的最近(約9845年から約13,027年前;
南米到達以降)に偶然に起こり、集団に広がったことが示唆されました。
アンデス人における比較的低いヘマトクリットの人々に見られる表現型は、チベット人のそれと類似していますが、アンデス人の変異がアミノ酸配列に起こった変
異であるのに対し、チベット人の変異はアミノ酸配列には対応しない遺伝子発現の制御に関わる領域における変異である点が異なっています。
すなわち、48,000年以上前にデニソワ人から受け継いだとされるチベット人のEPAS1の変異と、比較的最近に起こって広がったアンデス人のEPAS1
の変異は全く異なるものですが、高地に順応するという、同じような表現型を呈するものとなったのです。
29.5 東アジアにおける適応的変異
第13回GWASで述べたようなゲノムワイドな解析により、東アジアでも地域特異的な強い自然選択の痕跡が検出された遺伝子がいくつか見つかっています。
(以下、木村亮介、日本生理人類学会誌 (2019) 24,87-91 を参照した)代
表的なものに、ABCC11、ADH1B、EDARがあります。ABCC11(Gly180Arg)はアポクリン腺分泌機能に影響を及ぼし、耳垢の乾湿型や体臭に
関連しています。Arg変異は東アジア人特有の乾いた耳垢や体臭の少なさに関与しています。ADH1B(His48Arg)は、第6回酵素で詳しく振れたよう
に、
アルコールからアセトアルデヒドへと分解する酵素の遺伝子で、東アジア人に特異的にみられるArg変異では、その反応速度を高める効果があります。最後の
EDAR(Val380Ala)は、毛、歯、爪、汗腺、皮膚腺など外胚葉由来の付属器の発生に関与する遺伝子で、東アジア特有のAla変異は、太い毛髪、
シャベル型切歯などに関与しています。
しかし、こうした変異がなぜ、自然選択を受けてきたのかはよくわかっていません。耳垢が乾いている、お酒に弱い、毛髪が太いことが、生存にどれだけ寄与す
るのだろう
かと思わざるを得ません。しかし、これらの遺伝子は、さまざまな形質に関与していることも知られており、それぞれ、もっとも身近な一つの形質だけに結
び付けて考えていることが、その困惑を生む原因となっているかもしれません。たとえば、EDARは上に掲げた表現型以外にも、顎の形態、耳たぶの形態、髭
の密度、エクリル汗腺の密度などにも関与していることがわかっています。そして、それらすべての表現型が適応的である必要はなく、これらのうちひとつが、強い選択圧を受
けてきたと考えることも可能なのです。
自然選択の正体を科学的根拠に基づいて特定することはなかなか難しい作業です。しかも、過去に大流行した感染症に耐性を示したことがその原因となっている
可能性もあるわけで、ますます困難を極めることになるやもしれません。しかし、それだけに、研究者にとっては、興味の湧いてくる話題でもあるのでしょう。
今回は雑学的な話題に終始しました。まだまだ話題はありますが、この辺で終わりにしたいと思います。従来、進化は少なくとも数百万年のレベルで起きる出来
事
と考えられていました。しかし上で掲げた例は、どれも数千年から1万年という期間で起きていることです。しかも、全員がある一つのタイプの遺伝子をもつように
なったというわけではなく、集団内に多型があるという話でもあります。ダーウィン的な進化の概念では適応度の高い遺伝子が集団を席捲するというイメージだった
のですが、多型を生み出すという側面も進化にあることをこうした話題は例示してみせてくれています。