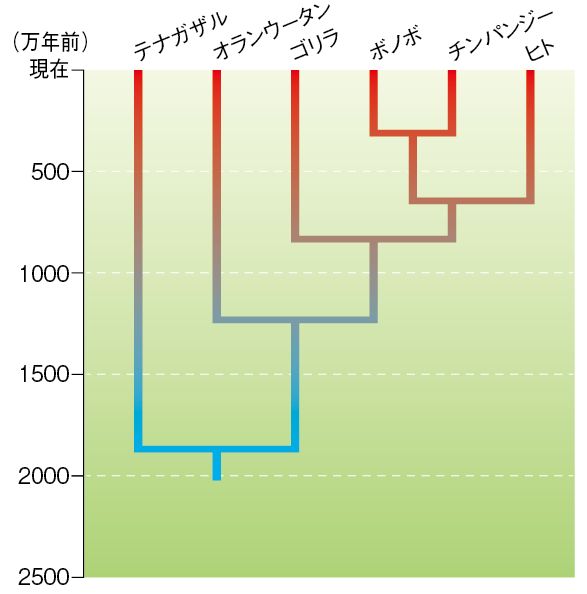
図28.1 ヒトと類人猿の系統樹
図の出典:https://www.zennoh-weekly.jp/wp/article/3594
「進化的にヒトに一番近いサルは?」という質問に、おそらく多くの方はチンパンジーと答えるでしょう。でも、私の世代では諸説あり、オランウータン説が有
力
であったように思います。多くの身体的特徴がヒトと一番よく似ているのがオランウータンだったからです。また、1932年、インドでラマピテクスと名づけられ
た類人猿の下顎の骨が発見されました。上顎骨と下顎骨の一部分しかない不完全なものでしたが、推定される歯列は放物線形で、類人猿の歯列はU字形であるのに対
して人類のそれは放物線を描いていることから、ラマピテクスは類人猿からヒトへとつながる生物、すなわちヒトの祖先種であると考えられました。さらに調査の結
果、ラマピテクスの生息年代は1400万年前と推定され、この時期がヒトと他の類人猿との分岐と考えられるようになりました。
ところが、前回お話しした分子時計の技術が開発され、現存する類人猿のDNAの塩基配列の比較が可能となると、意外な結果が出てきたのです(図1)。ヒト
に
一番近い(すなわちDNAの塩基配列が似ている)のはチンパンジー、次にゴリラで、オランウータンは進化的にはもう少し遠縁の存在というのです。しかも、ヒト
とチンパンジーが分岐したのは500万年前。ラマピテクスで推定された年代よりはるかに最近分岐したというのです。
その頃、分子時計の理論はまだすべての科学者が認めるほどには認知されていませんでしたから、この結果には多くの反論が寄せられました。日本人にはわかり に くい感情かもしれませんが、サルがヒトの祖先であることを受け入れ難い人たちも多く、たとえ受け入れたとしても、たった500万年前(進化的にはつい最近)サ ルから分岐したということは、われわれはかなりチンパンジーに近い存在だということになり、そうした事実を認めたくないという思いなどもあって、この分子時計 による推定はすんなりと受け入れられませんでした。特に、ラマピテクスの化石は強力な反論となりました。ところが、1982年、ラマピテクスの完全な頭骨が新 たに発見されると、歯列の形の推定が誤っていた事がわかり、オランウータンの系統であることが明らかになり、反論の根拠を失います。さらに、分子時計の正当性 がさまざまな事例で証明されるようになり、500万年前(現在の推定では600万年から700万年前と少し遡っています)にチンパンジーと分岐したということ が認められるようになりました。
ちなみに、上記の分子時計では、DNAの一部の比較による推定ですが、2000年代にはいると、ヒトおよびチンパンジーの全ゲノムが解読され(およそ30 億 塩基対からなるDNA全体の塩基配列がわかることをこう表現します)、両者の違いはたった1.23%であることがわかりました。ヒトどうしを比較しても平均 0.1%程度の違いがあるとされていますから、いかに小さな値かがわかります。
(注) 1980年代初め頃、ヒト上科の分類は、テナガザル科(テナガザル)、オランウータン科 (オランウータン、ゴリラ、チンパンジー)、ヒト科(ホモ・サ ピエンス)となっていました。分子時計の結果を受け、この分類は変更され、テナガザル科(テナガザル)、オランウータン科(オランウータン)、ヒト科(ゴリラ 属:ゴリラ、チンパンジー属:チンパンジー、ヒト属:ホモ・サピエンス)と変わり、当時、この改訂を「ヒトになったチンパンジー」と報じた新聞もありました。 さらに、 2000年代のゲノム解読を受け、チンパンジーをヒト属に分類すべきとする提言もなされています。
1980年代半ば、分子時計がかなり認知されるようになったころ、できるだけ多くの民族を含む147人のミトコンドリアDNAの塩基配列を比較し、その
147人の系統樹を推定するという研究が行われました。ミトコンドリアDNAとは、核ではなく、ミトコンドリアという細胞内器官に存在するDNAのことで、核
にあるのが30億塩基対のDNAが2組であるのに対して、2万塩基弱という小さなDNAです。しかも核にあるDNAと大きく異なる点に、母性遺伝するという特
徴があります。
われわれは男性、女性を問わず、ミトコンドリアDNAをもっていますが、それらはすべて母親から受け継いだものです。精子の中にもミトコンドリアDNAが 含 まれているのですが、受精後すべて分解されてしまいます。したがって、男性は、いまもっているミトコンドリアDNAを子孫に伝えることはありません。見方を変 えると、皆さんのもっているミトコンドリアDNAは母親から譲り受けたものであり、母親はその母親から譲り受けたものであり、・・・。すなわち、母系を通じて 伝わってきたものです。
これに対して、核にあるDNAは、Y染色体を除き、父親と母親それぞれからもらったものがあり、たとえば母親からもらったDNAは、母
親はその父親からもらったかもしれないし、その母親からもらったかもしれません。そうして、祖先を尋ねようとすると、とても複雑な状況となることがわかりま
す。ちなみに、Y染色体にあるDNAは、男性しかもちませんし、当然父親からもらったもので、父親も、その父親からもらっていますので、こちらは父系を通じて
伝わったものです。ミトコンドリアDNA同様、その祖先をすっきりたどることができます。ただし、Y染色体のDNAはおよそ5千万塩基対もあり、ミトコンドリ
アDNAに比べ格段に大きいため、ミトコンドリアDNAの解析に後れを取りましたが、最終的には、それぞれの解析から、現生人類の系統樹が描かれました。
ミトコンドリアDNAとY染色体DNAの解析から描かれた系統樹は図2のようになりました(ここでは、最初に解析した研究者の研究結果ではなく、その後多 くの 研究者が追試した結果を示すことにします)。両者はほぼ同じような結果が導かれています。この系統樹からは、現生人類の共通祖先は10数万年前に遡ることができ ます。そこから分岐が起こりますが、その分岐の仕方を見ると、6万年前にアフリカからアジアへ、4万年前にアジアからヨーロッパへ、そして1万数千年前にアジ アからアメリカ大陸へと分岐が起きているのが分かります。すなわち、現生人類はすべてアフリカ起源で、その後、アジア、ヨーロッパ、そしてアメリカ大陸へと拡 散していったことがうかがえます。
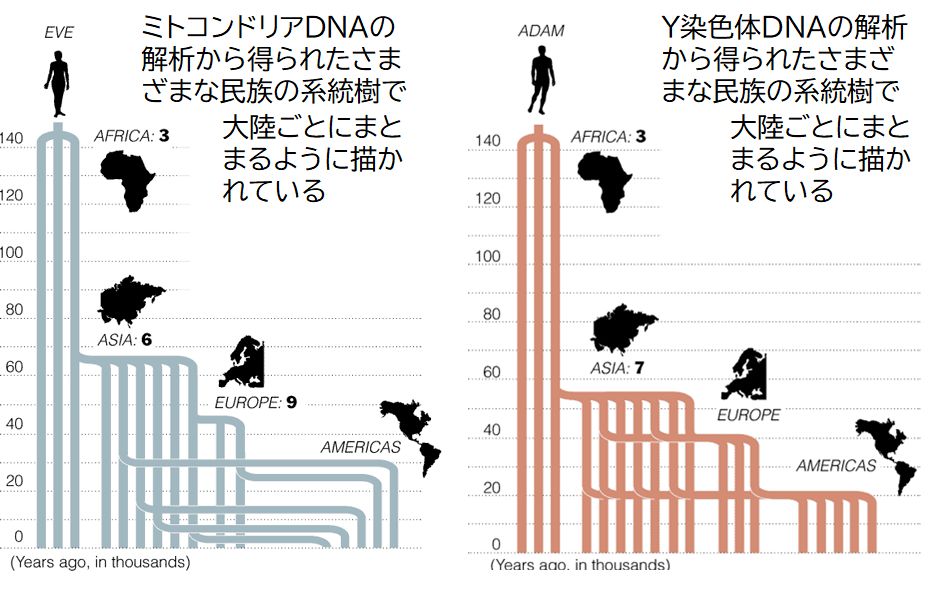
図28.2 ミトコンドリアDNAおよびY染色体DNAに基づいて作られたヒトの系統樹
系統樹のはじめは、ミトコンドリア・イブ、Y染色体アダムと呼ばれた。
図の出典:NY Times (May 2, 2000) "The Human Family Tree: 10 Adams and 18
Eves"
(注) 図28.2にミトコンドリア・イブ、Y染色体アダムという言葉が見られます。ミトコンドリアDNAを調べられた人の母系を遡ると、いつか全員の共通の
祖先、
すなわち一人の女性にたどり着きます。したがって、現生人類はその女性から始まったと考えられ、それをミトコンドリア・イブとよびました。Y染色体についても
同様な論理から、現生人類最初の男性としてY染色体アダムが存在するとされました。
しかし、この論理には重大な誤解があります。確かに、現代生きている人全員のミトコンドリアの共
通の祖先がいることは論理的に導かれます。しかし、われわ れ
の核にも、それとは比較にならない量のDNAがあります。たとえば、その中の2本ある第1染色体は、その1本を父親から、もう1本を母親から受け継いでいます
が、その父親から譲り受けた第1染色体を遡っていくことを考えてみましょう。それは母系でもなく、父系でもありませんから、複雑な形で遡ることなりますが、そ
れでもミトコンドリアDNAと同様、現代を生きている人全員の第1染色体の共通の祖先が存在することになります。しかしそれは、ミトコンドリア・イブとは異な
る人になるはずです。さらに複雑なことには、核にある染色体は組み換えを起こしますから、第1染色体がそのままの形で遺伝するわけではありません。したがっ
て、第1染色体を細かく領域に分け、領域ごとに共通の祖先へと遡る必要があります。そして、たどり着いた共通祖先は、領域ごとにすべて異なる人となります(領
域によっては同じ人となっても構いません)。
閑話休題。こうした解析から、現生人類はすべて十数万年前アフリカにいたあるグループの子孫で、その後世界に拡散していったことが推定されました。その結果
をまとめたのが図28.3です。日本には3〜4万年前にたどり着いたと推定されています。
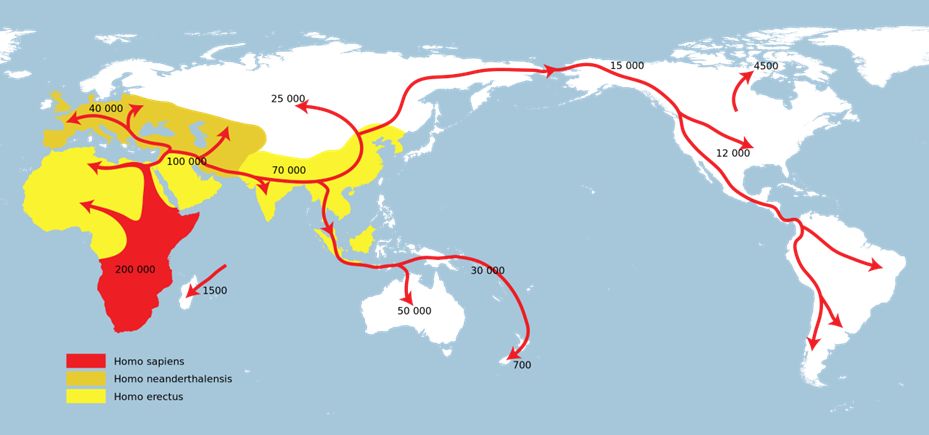
図28.3 推定される現生人類の世界拡散の経路とその時期
図の出典:Early human migrations @Wikipedia
ところで、チンパンジーとヒトが分岐したのは600〜700万年前でした。160〜170万年前にいたとされるジャワ原人、70万年前にいた北京原人、約
40万年前に出現し、2万数千年前に絶滅したとみられるネアンデルタール人、いずれもこの現生人類の祖先が出アフリカする前にアフリカ以外の地域に住んでいた
人類です。日本でも、およそ12万年前の前期旧石器時代の遺跡が島根県で発掘されています。これはどういうことでしょうか。
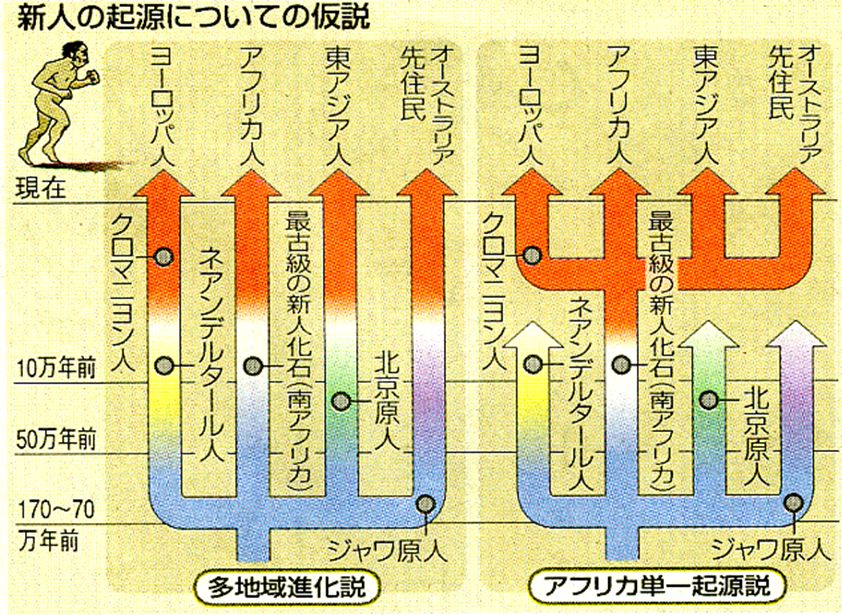
私たち現生人類(新人)の起源に関しては、化石などのデータを基に、世界各地の原人がそれぞれの地域で新人へと進化したという「多地域進化説」がありまし た。 しかし、上記の結果は、ジャワ原人、北京原人、ネアンデルタール人、そして日本の前期旧石器時代にいた人類(彼らは旧人とよばれる)は絶滅したことを示唆して います。十数万年前にアフリカにいた現生人類の祖先が、世界中に拡散して旧人の子孫に取って代わったと解釈せざるを得ず、これを「アフリカ単一起源説」といい ます。ただ、多地域進化説でも、その起源はアフリカにあると考えられており、170万年前ころに出アフリカを果たしたと推定されていますので、アフリカ単一起 源説という命名は適切かどうかわかりません。したがって、2つの説の比較では、共通祖先にたどり着くまでの時間に大きな違いがあり、現生人類の遺伝的な違いは 「アフリカ単一起源説」の方が「多地域進化説」よりはるかに小さいことが重要な点となります。
新人類がアフリカを出て世界中に広がり始めると、それまでユーラシア大陸に住んでいた同じホモ属のネアンデルタール人などの旧人類は絶滅へと追いやられまし
た。どのような形で旧人類が競争に負け、新参のホモ・サピエンスに取って代わられたのかは今後の研究が待たれますが、少なくとも、ネアンデルタール人と現生人
類の祖先が共存していた時代があることがわかっています。そのとき、新人類は旧人類とは交わらず、ネアンデルタール人の遺伝子を次世代に伝える混血の子どもは
生まれなかったと考えられてきました。
ところが、ここ約10年の化石人類の発見ラッシュと遺伝学的研究の発展によって、この定説は大きく書き換えられることになったのです。その端緒となったの は、2010年、ネアンデルタール人の全ゲノムが解読され、現生人類の中にネアンデルタール人特有の遺伝子が存在することが明らかになったことでした。混血は 起 こっていたのです。ただし、アフリカ人の中には見つかりませんでした。したがって、出アフリカを果たし後、交配が行われたと推定されます。また、今日の非アフ リカ系の人々のゲノム(全遺伝情報)の最大3%がネアンデルタール人由来と推定されています。さらに、人によってそれぞれネアンデルタール人由来の異なる DNA断片を持っているため、現生人類が受け継いだネアンデルタール人の遺伝情報の総和は少なくとも20%にはなるだろうと推定されています。
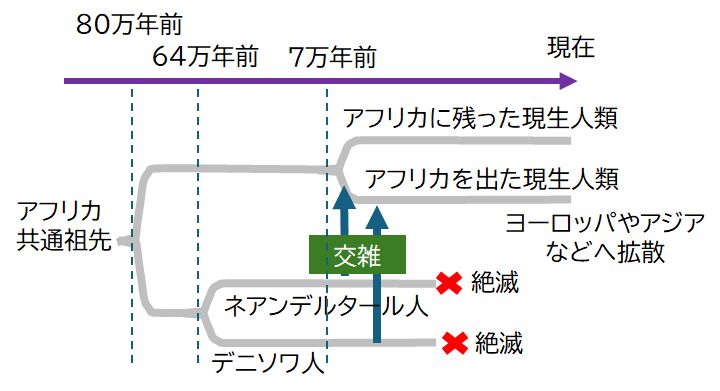
(注)ネアンデルタール人にゲノムを解読したスバンテ・ペーボは、2022年、ノーベル医学・生理 学賞を受賞しました。
ホモ・サピエンスとの混血があった旧人類はネアンデルタール人だけではありませんでした。2008年に発見されたデニソワ人(シベリアのデニソワ洞穴で見
つ
かった4万年ほど前の謎めいた指の骨から回収されたDNAによって特定された人類集団)も、オーストラリア北東の島々に住むメラネシア人は、ゲノムの4〜6%
がデニソワ人固有のものと一致しており、現生人類の先祖との間に混血があったことがわかってきました。デニソワ洞穴には石器などから12万5千年前には住んで
いたことがわかっており、そうした点からも現生人類の祖先と共存していたことがわかります。なお、約80万年前にデニソワ人とネアンデルタール人の祖先が現生
人類と分岐し,約64万年前にネアンデルタール人とデニソワ人が分岐したと推定されています。
分子時計は、お互いのDNAを比較し、違いを数えることから分岐からの時間を推定するのが基本です。しかし、ヒトどうしのように、その数が小さい場合誤差 も 大きくなるため、慎重に取り扱う必要があります。そこで、DNAの変異をより詳細に比較するため、変異をいくつかのタイプに分け、そのタイプの構成比から、ヒ トの進化を解析する方法が提案されました。そうした変異のタイプをハプロタイプ、同じハプロタイプをもつ人々をハプログループとよびます。
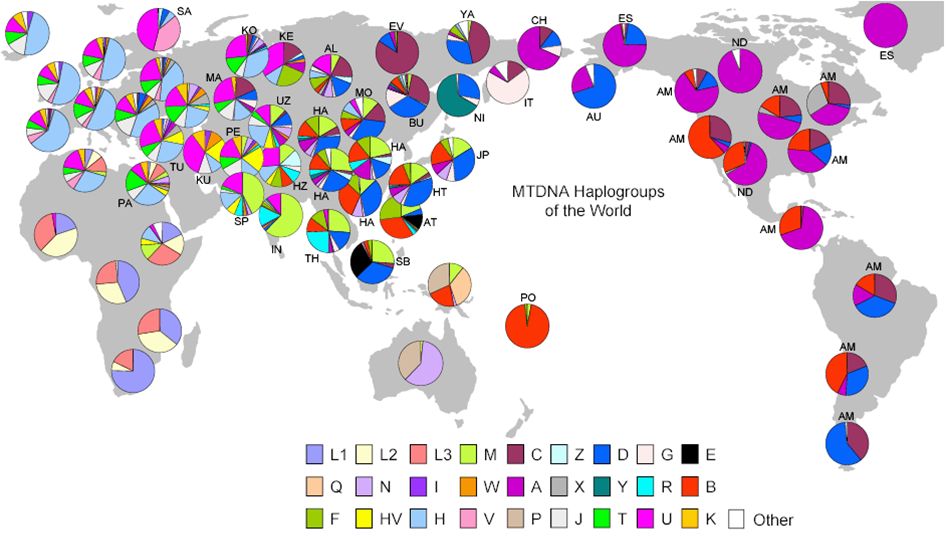
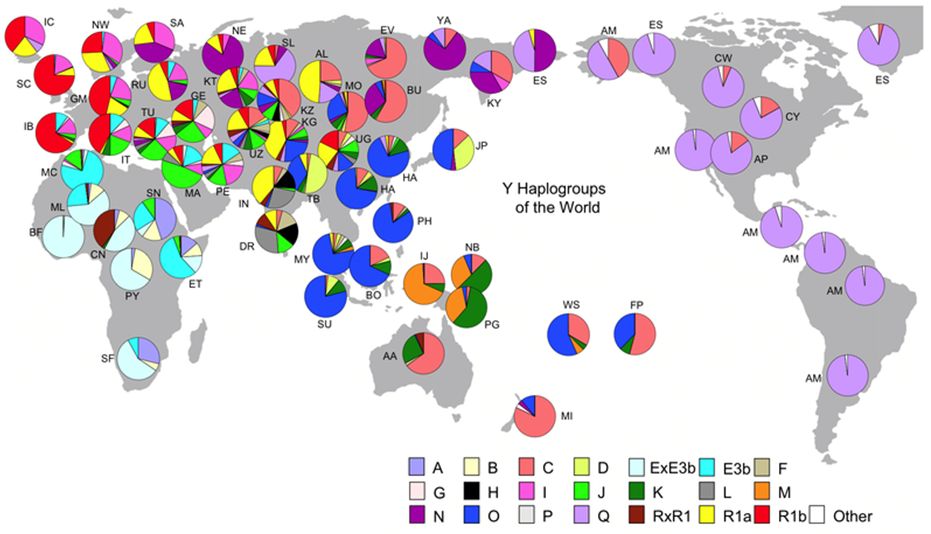
図28.6上にミトコンドリアDNAの、そして下にY染色体DNAのハプログループの分布を示しました。同じ色が同じハプロタイプをもつハプログループの割
合を 示
しており、アフリカから世界へと現生人類が拡散していった様子がこうした図からも読み取ることができます。また、それぞれの地域で複数のタイプが共存している
のが読み取れ、そこから人々の交流の歴史をいろいろと考えさせてくれます。
ところで、ミトコンドリアDNA(母系)とY染色体DNA(父系)と見比べて何か不思議に感じたことはありませんか。東アジアをみると、ミトコンドリア
DNA
の円グラフは日本も含め、互いによく似ているのに対して、Y染色体では、日本と中国大陸では大きく異なっています。日本はむしろチベットと近いといえます。なぜ
なんでしょうか?ヒントは女性と男性では子孫の残し方に差があることです。
女性が生涯にもうける子供の数はある程度制限されますが、男性の場合、はるかに多くの子孫を残す可能性があります(注:逆に言えば、全く残せない男性もい
る)。また、女性は相対的にあまり移住せず、その地域に根を下ろして生活しているのに対して、男性は移動範囲が大きく、とりわけ領土拡大などの戦争に参加し勝
利すれば、その遠征先で子孫を残すことも可能です。こうした違いが、ミトコンドリアとY染色体のDNAのハプログループの分布の差となって表れたと考えられま
す。
Y染色体のD2(図28.6の黄緑)の分布は、もともと北東アジアに広く分布していましたが、その後中国を中心とした地域で勢力を伸ばしたハプログループ
Oの系統 (図28.6の青;漢族)によって周辺に押しやられてしまった結果、こうした分布になったと思われます。
しかし、日本ではD2がそのまま残り(縄文系の人々)、縄文から弥生への移行期、渡来人(O;弥生系の人々)の流入が平和的に行われた姿が浮かび上がって
き ま
す。もし、渡来人が縄文時代から続いた在来社会を武力によって征服したのであれば、その時点でハプログループD2は、著しく頻度を減少させたでしょう。
10.3の結果から、ハプログループ、とりわけY染色体のハプログループの分布から人の移動の歴史が読み取れることが分かりました。そこで、図28.7 に、メソ ポ タミア文明で始まった農耕文化がどのようにヨーロッパで広まっていったかを示した地図と、Y染色体のJ2というハプロタイプの現在の頻度分布を並べてみまし た。ハプロタイプの頻度分布は、特に適応度に影響せず、自然選択が起こらなければ、頻度の高いところが発祥の地で、そこから拡散していった様子を表していると 考えられています。図28.7の2つの分布がよく似ているのは、農耕文化が口承による伝播ではなく、人の移動によるものであることを表していると捉えられてい ます。
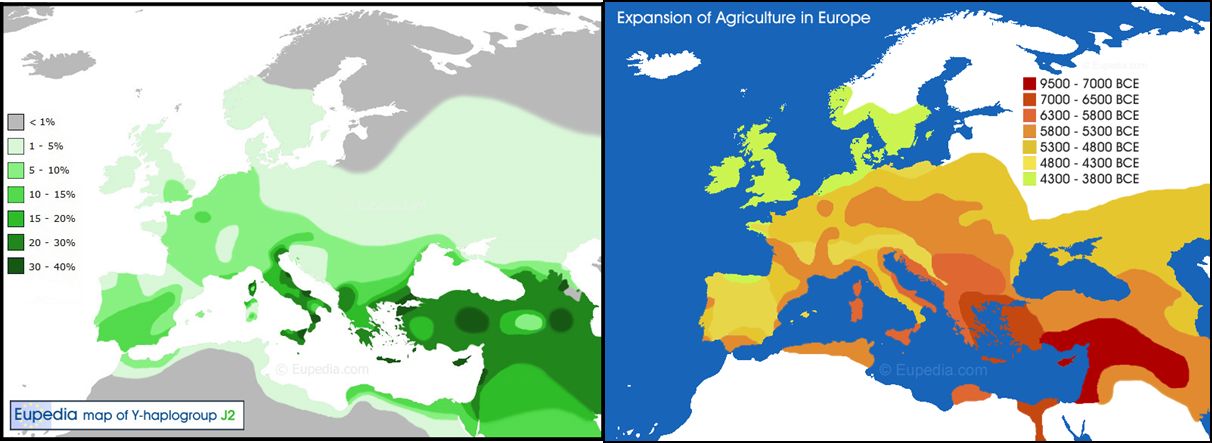
こうした分野にも、DNAの解析は利用されるようになっているのです。
![]()
外部リンク 縄文人と弥生人で分けられない「日
本人のルーツ」 篠田 謙一 : 国立科学博物館
東洋経済ONLINE 2024/04/14