�@�O��́A�Ɖu�̑S�̑���b�����ƂȂ��A�R�̂ɓ������Ęb���n�߂܂����B����́A�Ɖu�̑S�̑������n���Ȃ���A�ʂ̘b��ւƂ͂����Ă��������Ǝv���܂��B����̍u�`���e �́A���̍u�`�S�̂̒��ł��ł����G�ł��B�����Řb���Ƃ��Ƃ͑S���قȂ�A���͂ɂ��Ă݂�Ǝ��ɓ���ȕ��͂ɂȂ��Ă��܂��܂����B��������Ȃ��p�ꂪ�p�o���A�� �̈��̃C���[�W�����グ��̂Ɏ��Ԃ�������A1��ǂݒʂ��������ł͓��ɓ����Ă��Ȃ���������܂���B�܂��A�����̈ꕔ������̍u�`�ł���悤�ɂ��� �b�������A�������Ă��Ȃ����Ƃ��A��������荢��ɂ��邩������܂���B���x���ǂݕԂ��Ȃ��痝���ɓw�߂Ă���������K���ł��B
�@�Ɖu�͑傫�����R�Ɖu�Ɗl���Ɖu�ɕ������܂��B���R�Ɖu�́A���������Ƃ��̏����Ή����s���܂��B�a���̂́A�����q�g�ɂ͂Ȃ��A�ۂȂ�ہA�E�C�� �X�Ȃ�E�C���X�ɋ��ʂ������q�����������Ă��邽�߁A�����̐H�זE�́A���������a���̂ɋ��ʂ�������F���ł��镪�q���זE�\�ʂ�����ɂ����Ă��܂��B�� �� ���Ɋ�Â����F�����@�ł��̂ŁA���Ȃ���ނ̕��q�ő�R�̎�ނ̍ۂ�E�C���X�ɑΉ����邱�Ƃ��ł��A�ۂ�E�C���X���̓��ł܂����܂葝�B���Ă��Ȃ������� ������ ���Ƃ��āA����Ȃ�̌��ʂ����܂��B���ہA�V�^�R���i�Ŗ��Ǐ�҂������̂́A���R�Ɖu�̒i�K�ŏ������������Ă��܂��Ă��邩��ł͂Ȃ����Ƃ������������� ��܂����B�������A�ЂƂ��уE�C���X�����B���ėʂ𑝂��ƁA��������ԑŐs�ɂ���ɂ́A���R�Ɖu�͕͗s���ł��B����ɁA�ő�̖��_�́A�זE�̒��ɓ��荞�� �� �E�C���X�ɂ͎���o���Ȃ����ƂŁA���ꂪ����I�Ȏ�_�ƂȂ��Ă��܂��B
�@�����������ԂɑΏ����邽�߁A���R�Ɖu�Ɉ��������l���Ɖu���N������܂��B���R�Ɖu�́A�l���Ɖu�ɕa���̂̏���`���A�N���̎�`�������܂��B�l���Ɖu�� �� ���́A����̕a���̂�F�����A������W���I�ɍU������\�͂������Ă��邱�Ƃł��B�܂��A�זE���ɐN�������E�C���X�������זE����Ƃ��j�A�����r�����邱�� ���ł��܂��B�������A�O�b�������悤�ɁA�����זE�͂��Ƃ��Ǝ����̍זE�ł��B���������āA����Ȏ��ȍזE���U���������ςȂ��ƂɂȂ�܂��B���Ƒ��� ���m�ɔ��f���邱�Ƃ��ɂ߂ďd�v�ȉۑ�ƂȂ�܂��B
�@�Ȃ��A���R�Ɖu�Ƃ����p��͈Ӗ����킩��ɂ����A�p��� innate immunity �����I�Ɖu��p�����������I�m�ŁA�ꕔ�̖{�ł͂��̗p����g���Ă��܂��B�������A���łɒ蒅���Ă���u���R�Ɖu�v�������ł��g�����Ƃɂ��܂��B�v���� �ɁA���R�Ɖu�Ƃ́A�a���̂ɂ��Đ��܂�Ȃ�������Ă�����𗘗p�����Ɖu�V�X�e���ł���A�l���Ɖu�́A����a���̂Ɋ����������Ƃŏ��߂Ă��̕a���̂̏����l �����A���̕a���̓��ٓI�ɏW���U�����s���V�X�e�����Ƃ������Ƃł��B
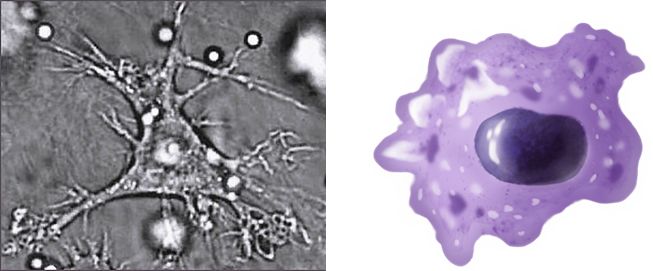
�@���R�Ɖu�n�ł́A����\�킸���ł��H�ׂ�H�זE�Ƃ���זE�����܂��B�}�N���t�@�[�W�A�D�����A����זE�ȂǂŁi�}24.1�j�A�����̍זE�̎��[�� �V �p���A �����ĐN�����Ă����a���̂ȂǁA����\�킸�H�ׂāA�������܂��B�������A�����Ă��鎩���̐���ȍזE�͐H�ׂ܂���B����ȍזE�ɂ� �gDon�ft eat me�h �̖ڈ��Ă��邩��ł��B
�@�H�זE�́A���ȍזE�R���̂��̂�H�ׂĂ����������܂��A�a���̂�H�ׂ�Ɗ��������܂��B����������ƁA�����\�́A�E�۔\�͂������A�x���i�T�C�g�J �C ���j����o���đ��̐H�זE�i�D�����A�}�N���t�@�[�W�j���ĂяW�߁A���ǂ������N�����܂��B���ǂƂ����ƁA���M�A�ɂ݁A���ȂǕs���ȏǏo�܂��̂ŁA�a���� �������N�����a�I�ȏǏ�Ǝv�������ł����A����ꎩ�g�̑̂��g����邽�߂ɂ��h��@�\�̈�тƂ��Ĉ����N���������̂Ȃ̂ł��B
�@�Ƃ���ŁA�ǂ�����Ď����̍זE�Ǝ����łȂ��זE��H�זE�͌������Ă���̂ł��傤���B������s���Ă���̂��A�p�^�[���F����e�̂Ƃ���^���p�N���ŁA���̒��ł��� �� �悭�m���Ă���̂��A�g���l��e�́iTLR; Toll-like receptor�j�Ƃ���^���p�N���ł��B1990�N��㔼�ɍŏ���TLR��������ƁA2000�N��ɂ͂����Ď����玟�ƐV����TLR��������A���݁A�q�g�ł� 10 ��ނ݂����Ă��܂��B �@�@

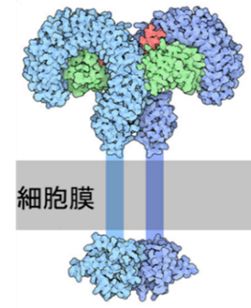
�@TLR�Ȃǂ̃p�^�[���F����e�̂́A�����͂������A�a���̂����������̕��q�ɑ���Z���T�[�ł��B���Ƃ��A�ۂ̍זE�ǁE�זE���̐����A�t���W�F �� ���i�ږу^���p�N���j�A�E�C���XRNA�A�E�C���XDNA�Ȃǂ������������̕��q�ł��i�}24.2 �Q�Ɓj�B�p�^�[���F����e�̂̓^���p�N���ł���A�e����q�ւƎp ���ꂽDNA�ɂ��̏����A����ɂ���Đ��������ꂽ���̂ł��B�O�b�������R�̂̂悤��DNA�̍č\���͍s���܂���B���ꂪ���R�Ɖu�i�����I�Ɖu�j�� ����R���ł��B
�@�Ȃ��A�H�זE�́A�p�^�[���F����e�̂��g���ĕa���̂��������A�a���̂�����I��I�ɐH�ׂĂ���킯�ł͂���܂���B����\�킸�H�ׂ���A�a���̂Ɣ��f���� ���犈�������A�x������o���܂��B
�@�}24.3�́A���R�Ɖu�Ɗl���Ɖu����Ȃ�Ɖu�V�X�e���̊T�v���܂Ƃ߂����̂ł��B���̐}���ŏI�I�ɗ����ł��邱�Ƃ��߂����Đ��������Ă����܂��B�������Ƃ����x���J�� �� ���Ă��b�����镔��������܂����A���G�ȃV�X�e���ł��̂ŁA�J��Ԃ����Ƃŗ�����[�߂Ă������������Ǝv���A�����ď璷�ȋL�q�ƂȂ��Ă��܂��B
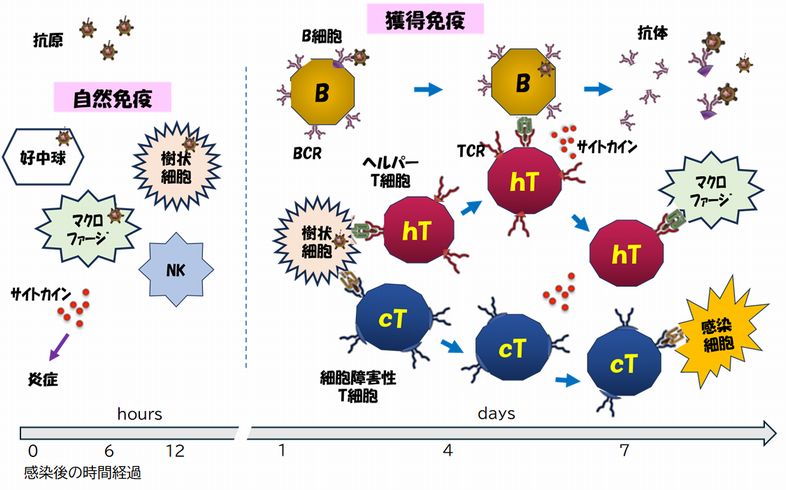
�@�܂��͐}24.3�ɕ`���ꂽ���Ƃ��܂Ƃ߂Đ������܂��B�ׂ��ȓ��e�͒ǂ��Đ������܂��̂ŁA�ŏ��͑S�̑��߂���x�ɓǂ݁A���̎����̍Ō�܂œǂݏI���
���Ƃ���ŁA������ �x�ǂݕԂ��Ă݂Ă��������B
�Z���R�Ɖu�i�����I�Ɖu�j �@
�@ �O������ۂ�E�C���X�Ȃǂ̍R�����N������ƁA�܂��A�}�N���t�@�[�W�A�D�����A����זE�Ȃǂ̐H�זE�̐H��p�ɂ�肱���r�����悤�Ɠw�߂܂��B
�A ����זE�́A�\�ʂ��邢�͓����ɂ���g���l��e�̂Ȃǂ̐����I�ɂ��p�^�[���F����e�̂ɂ��A�߂炦�����̂��a���̗R�����q�������ǂ������`�F�b�N���܂��B�a���̗R�� �̎� ��זE���������q�́A�Ɖu�A�W���o���g�ƌĂ�܂����A�ۂ̍זE�ǁE�זE�������A�ږу^���p�N���A�E�C���XRNA�A�E�C���XDNA�Ȃǂ�����܂��B����זE�� �ߊl�������̂̂Ȃ��ɂ����������q�̑��݂��m�F����ƁA�T�C�g�J�C������o���ĉ��ǂ������N�����悤�����A����זE���g������ɂ�芈�������܂��B�B ��������������זE�́A�H�ׂ��a���̂̃^���p�N�������A���̒f�Ђ�MHC�Ƃ��鎩�g�̃^���p�N���ɏ悹�A�זE�\�ʂŊl���Ɖu�n��T�זE�ɒ��܂��i�R���j�B ����ɂ��A�l���Ɖu�n���N�����܂��B �Ȃ��AMHC�ɂ̓N���XI�ƃN���XII������AT�זE�ɂ̓w���p�[T�זE�ƃL���[T�זE�i�זE��Q��T�זE�j������܂����A�����̈Ⴂ���܂߂��ڍׂ́A�ȉ��ɏq�ׂ邱�� �ɂ��܂��B
�Z�l���Ɖu
�@�l���Ɖu�̓����́A����̍ۂ�E�C���X���W���I�ɍU�����邱�Ƃł��B�����ł͂��̎�v�����o�[�ł���3��ނ̖Ɖu�זE�AB�זE�A�w���p�[T�זE�A�L���[T�זE�ɂ��� ���b�����܂��B �@
�@ B�זE�͍R�̂��Y������Ɖu�זE�ŁA�O��̍u�`�ł��łɂ��b�����܂������A�����ƕ��K���Ă����܂��傤�B B�זE�́A���̍זE�\�ʂ�B�זE��e�̂Ƃ����^���p�N���������Ă��܂��BB�זE��e�́iBCR: B-cell receptor�j�́AB�זE���Ƃɂ��̍R���������ʂ̌`���قȂ�A���牭��ނ͂���Ƃ���܂����A���ȗR���̃^���p�N���Ɍ���������̂͂��炩���ߔr������Ă��邽�߁A �a���̂Ȃǂ� �O���̃^���p�N���ɂ̂������邱�Ƃ��ł��܂��i���邢�́AB�זE��e�̂������������̂͊O���̃^���p�N���ł���Ɗm�F�ł��܂��j�B�Ȃ��A������B�זE��e�̂̌������ʂ� �O���^���p�N���̂���̈�i�G�s�g�[�v�j�̌`�����ƌ����̊W�ɂ���� ���̂݉\�ł��B �@�@
�@B�זE��e�̂��O���^���p�N���ƌ�������ƁAB�זE�͂�����זE���Ɏ�荞�݁A�������A���̒f�Ёi�R���y�v�`�h�j��MHC�N���XII�Ƃ����^���p�N���ɏ� �� �� [�R���y�v�`�h�{MHC�N���XII] �����̂��`�����A�זE�\�ʂŒ��܂��B���鑊��̓w���p�[T�זE�ŁA���L�A�Ő������銈�������ꂽ�w���p�[T�זE���T�זE��e�̂ƁA����B�זE��ɒ��ꂽ [�R���y�v�`�h�{MHC�N���XII] �����̂��A���ƌ����̊W�Ō��������B�זE�͊��S�Ɋ���������AB�זE��e�̂��R�̂Ƃ��čזE�O�֕��債�܂��B
�@B�זE��e�̂́A�������Ȑ����ɂ͌������܂���̂ŁA�����A���ȗR���̃y�v�`�h����邱�Ƃ͂���܂���B�������A�����������̂��{���Ɏ��ȗR���̂��� �łȂ� �����A���������ꂽ�w���p�[T�זE��T�זE��e�̂�B�זE������[�R���y�v�`�h�{MHC�N���XII]�����̂Ƃ��������邩�ǂ����ɂ��ēx�`�F�b�N���A�� �n�t������̂ł��B �@
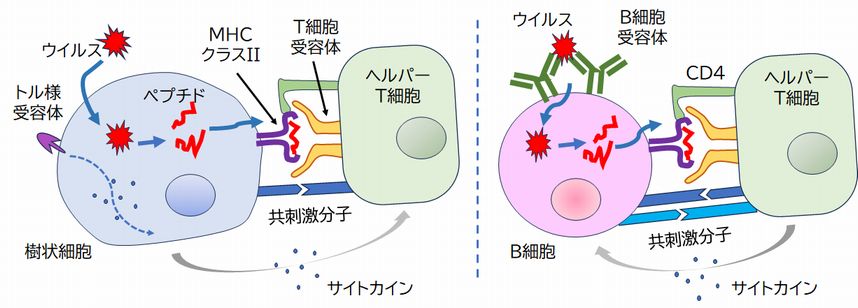
�A �w���p�[T�זE�́A���̍זE�\�ʂ�T�זE��e�́iTCR: T-cell receptor�j�Ƃ����^���p�N���������Ă��܂��BT�זE��e�̂̌������ʂ́A�R�̂Ɠ��l�A���牭��ނ̌`������A��ŏq�ׂ����R�Ɖu�n�ŕa���̂��ÐH���A���������ꂽ �� ��זE��� [�R���y�v�`�h�{MHC�N���XII]�����̂������ƁA���̂����̂ǂꂩ���������܂��B���̌����ɂ��A���̃w���p�[T�זE�͊��������A��L�@�ŏq�ׂ��悤�ɁAB�זE �� ���� ���̎菕�������܂��i���ꂪ�w���p�[�̗R���ł��j�B�w���p�[T�זE������זE���[�R���y�v�`�h�{MHC�N���XII]�����̂ɂ��AB�זE���[�R���y�v�` �h�{MHC�N���XII]�����̂ɂ����������Ƃ������Ƃ́A���҂������R���y�v�`�h������̂��̂ł��邱�Ƃ�ۏ��܂��B����́A����זE�̃p�^�[���F���� �e�̂ɂ��ٕ��̔F��ƁAB�זE��B�זE��e�̂ɂ��ٕ��̔F��Ƃ���v�����Ƃ������Ƃł���A���҂ɂ��_�u���`�F�b�N�ňٕ��Ɗm�F�������Ƃ��Ӗ����܂��B �@
�@�܂��A�w���p�[T�זE�́A�}�N���t�@�[�W�̊������ɂ���^���A�����ÐH�̓x���������߂܂��B �@
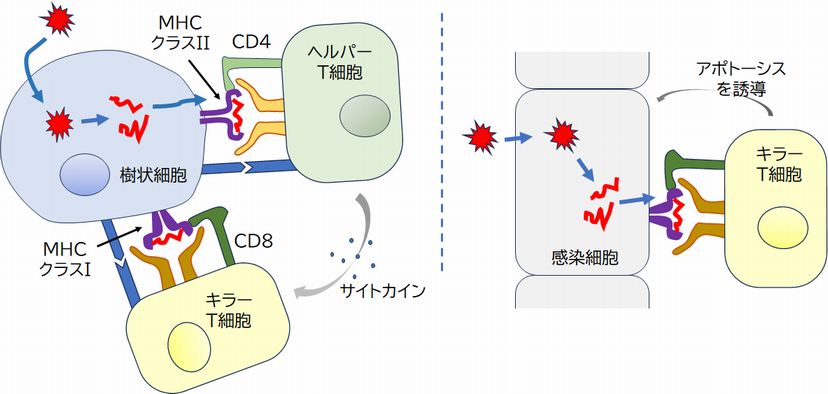
�B �L���[T�זE�i�זE��Q��T�זE�GCTL: Cytotoxic T lymphocyte�j�����̍זE�\�ʂ�T�זE��e�̂Ƃ����^���p�N���������Ă��܂��B�w���p�[T�זE��e�̂Ɠ��l�A���R�Ɖu�n�ŕa���̂��ÐH���A���������ꂽ����זE�� �� [�R���y�v�`�h�{MHC�N���XI]�����̂������ƁA���牭��ނ���T�זE��e�̂̂ǂꂩ���������A��������Ɗ��������܂��B�������A�����Œ��ӂ��Ȃ� �Ă͂Ȃ�Ȃ����Ƃ́A�R���y�v�`�h������Ă���̂́A�w���p�[T�זE�ł̓N���XII�ł������̂ɑ��āA�L���[T�זE�ł̓N���XI�ł��邱�Ƃł��B
�@����A��ʂ̍זE�ł��A�E�C���X���N�����A���B���邽�߂ɐ���ɃE�C���X�̃^���p�N�����זE���ō��������悤�ɂȂ�ƁA���̃^���p�N���̒f�Ђ�MHC�N �� �XI�ɏ悹�A�זE�\�ʂŒ���悤�ɂȂ�܂��B�����ŁA���������ꂽ�L���[T�זE�̂���T�זE��e�̂��A������ʍזE�̕\�ʂɂ��� [�R���y�v�`�h�{MHC�N���XI]�����̂Ɍ�������A���̃y�v�`�h�͎���זE���������̂Ɠ����ł��邱�Ƃ��m�F�ł��A���̍זE���E�C���X�Ɋ������Ă���Ɣ��f�o���� �킯�ł��B�����āA�L���[T�זE�͂��̊����������ȍזE�ɃA�|�g�[�V�X��U�����A���̊����זE����Ƃ������ɐ��ރE�C���X�����ł����܂��B �@�@ �@
�@�Ȃ��A�̉t���V����R�����R�̂ɂ���Ĕr������Ɖu��̉t���Ɖu�i���邢�͉t���Ɖu�j�A�זE�ɐN�������R���������זE����Ƃ��L���[T�זE�ɂ���Ĕr ������Ɖu���זE���Ɖu�Ƃ�т܂��B �@�@
�@ ���̖h��@�\�ɂ����ẮA���ȂƔȂ̔F�����ŏd�v�ۑ�ł��B���̔��f�����ƁA����Ȏ����̍זE���U�����邱�ƂɂȂ�܂��B���R�Ɖu�Ɗl���Ɖu�Ƃ��������āi�_�u�� �`�F�b�N�ɂ��j�A���̌��ɂ߂̊m�x�����߂Ă��邱�Ƃɒ��ӂ��Ȃ���ǂݐi�߂Ă��������B
�@�ȉ��ł́A�R�������T�זE��e�̂�[�y�v�`�h�{MHC]�����̂̊W�ɂ��āA����ɏڂ����������܂��B
�@��L�̌J��Ԃ��ɂȂ�܂����AT�זE�̓����������ƕ��K���܂��B
�@�Ɖu�����̕W�I�������R���Ƃ����܂��B�ہA�E�C���X�Ȃǂ̊O�����ٕ������łȂ��A����זE�̂悤�Ȉٕ����������ȗR���̍זE�Ȃǂ��܂܂�܂��B����זE �͑̂̋��X�܂ŒT�����Ă܂��A�������R����߂��ĐH�ׁA���̃^���p�N�������A�f�Љ����܂��B���`�\���̃A�~�m�_����Ȃ�^���p�N���̒f�Ђ��y�v�` �h�Ƃ�т܂����A���ɂ��̏ꍇ�͍R���̃^���p�N���̃y�v�`�h�Ȃ̂ŁA�R���y�v�`�h�Ƃ�т܂��B
�@�����̍R���y�v�`�h�́A��v�g�D�K�����R�� major histocompatibility complex (MHC) �Ƃ���^���p�N�����q�ɑg�ݍ��܂�A�זE�\�ʂɔz�u����܂��B��v�g�D�K�����R���Ƃ������̂��̂������O�̗R���͎��b�����܂��B�܂��A���Ƀq�g��MHC�� HLA�iHuman Leukocyte Antigen���q�g�������R���j�Ƃ����܂����A�����ł�MHC�̌Ăі���p���邱�Ƃɂ��܂��B
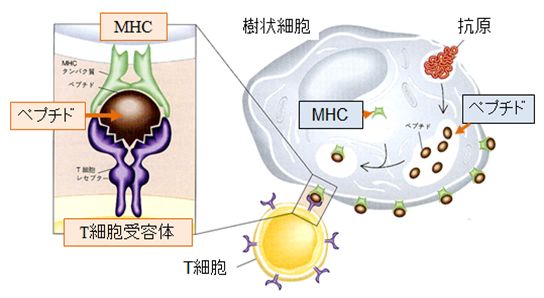
�@����AT�זE�ƌĂ�锒�����́A�X�ɈقȂ����������ʂ�����e�̃^���p�N���AT�זE��e�̂����̕\�ʂɂ����A�R���זE������[�R���y�v�` �h�{MHC]�����̂ƌ��ƌ����̊W������T�זE��e�̂������A����Ɍ������邱�Ƃ��ł��܂��B�����āA��������T�זE��e�̂�����T�זE�͊���������A �� ����J�n���܂��B�Ȃ��AT�זE�ɂ͍זE��Q��T�זE�i�L���[T�זE�j�ƃw���p�[T�זE������܂��B�L���[T�זE�́A[�R���y�v�`�h�{MHC]�����̂����̕\�� �ɒ��銴���זE��F�����A����ɃA�|�g�[�V�X��U�����A�j�邱�ƂōזE���ɐN�������E�C���X�����ł����܂��B�w���p�[T�זE�́AB�זE��}�N���t�@�[ �W�����������܂��B���������ꂽB�זE�̓v���Y�}�זE�ƂȂ�AB�זE��e�̂�ɎY�����A�R�̂Ƃ��ĕ��債�܂��B�܂��A���������ꂽ�}�N���t�@�[�W���ÐH�� �x���������߂܂��B
�@T�זE�̓����̑�܂��ȗ���͈ȏ�ł����A�}24.5���Q�Ƃ��Ȃ���AMHC�ɂ��R���ɂ��Ă����������̏ڍׂ����Ă����܂��傤�B
�@����זE�́A���ȗR���̍זE�A�a���̂��킸�H�ׁA�\������^���p�N�������A���̒f�Ёi�y�v�`�h�j��MHC�N���XI�����MHC�N���XII���q�Ɍ��� ���A�זE�\�ʂɒ��܂��B����A�a���̂����Ȃ��ł́A����זE�͐}24.5�iA�j�̂悤�Ȍ`������Ă���A���ȍזE�R���̃^���p�N���f�Ёi���ȃy�v�` �h�j�̂� MHC�Ɍ������čזE�\�ʂɒ��Ă��܂��B�������A����זE���a���̂�H�ׁA���g�����g���l��e�̂Ȃǂ̃p�^�[���F����e�̂�������ٕ��Ɣ��f����Ɗ��� �����A�iB�j�̂悤�Ɏp��ς��i���̌`������זE�Ƃ���R���ł��j�A���̕\�ʐς��L���A�a���̗R���̃^���p�N���f�Ђ�MHC�N���XI�����MHC�N���X II���q�Ɍ������A����悤�ɂȂ�܂��B�Ȃ��A���̂Ƃ���̍R�����瑽�l�Ȏ�ނ̃y�v�`�h�����ݏo����܂����AMHC�ƌ����ł�����̂���������� ���B
�@����AB�זE�́A���̕\�ʂɂ���B�זE��e�̂ɍR������������ƁA������זE���Ɏ�荞��ŕ������AMHC�N���XII���q�ɏ悹�čזE�\�ʂŒ��܂� �i�}24.5(C)�j�BB�זE��e�̂́A�������Ȑ����ɂ͌������܂���̂ŁA���ȗR���̃y�v�`�h����邱�Ƃ͂���܂���B�������A�����������̂��{���Ɏ� �ȗR���̂� �̂łȂ����A����זE�Ƃ̌����ɂ�芈�������ꂽ�w���p�[T�זE��T�זE��e�̂�B�זE������[�R���y�v�`�h�{MHC�N���XII]�����̂Ƃ��������邩�� �����ɂ��ēx�`�F�b�N���s���܂��B�����w���p�[T�זE���A���������ꂽ����זE�̒���[�R���y�v�`�h�{MHC�N���XII]�����̂ɂ��AB�זE������ [�R���y�v�`�h�{MHC�N���XII]�����̂ɂ��A���ƌ����̊W�Ō������邱�ƂŁA�R���y�v�`�h������̂��̂ł��邱�Ƃ��F�肳��܂��B
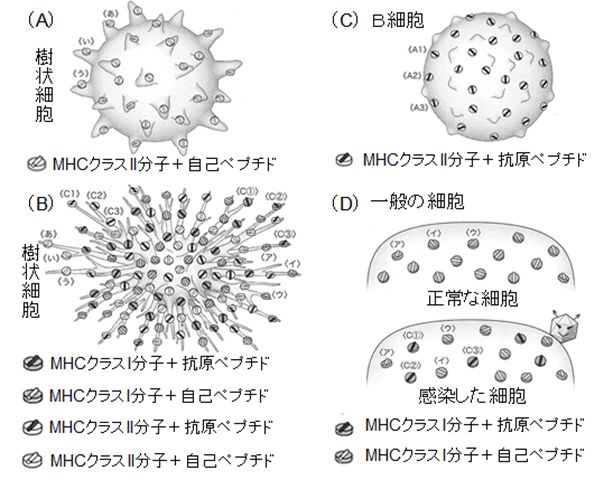
�@��ʂ̍זE���A���i����R�����s���Ă��܂��i�}24.5(D)�j�B�����͍̂זE���ō������ꂽ�^���p�N���̒f�ЁA���Ȃ킿���ȃy�v�`�h�ŁA MHC�N���X I���q�ɏ悹�čs���� ���B�זE���������Ă��Ȃ���A[���ȃy�v�`�h�{MHC�N���XI] �����̂���������܂��B��������� [�R���y�v�`�h�{MHC�N���XI]�����̂������悤�ɂȂ�܂��B�L���[T�זE�͂��������זE��F�����A�U������̂ł��B
�@�Ȃ��A��ʂ̍זE���p�^�[���F����e�̂������A�a���̂Ɋ�������ƌx���i�T�C�g�J�C���j�債�܂��B����ɗU������ăL���[T�זE�������זE�ɏW�� ���Ă��܂��B
�@�����ŁAMHC�N���XI�ƃN���XII�̈Ⴂ�ɂ��Đ����������Ǝv���܂��BMHC�N���XI�͑̒��̍זE�������A�זE���ō������ꂽ�^���p�N�����ꕔ�������č��ꂽ�y�v �`�h���悹�܂��B�E�C���X�́A��������ƁA���̍זE���̋@�\��q���A���B���邽�߂ɐ���Ƀ^���p�N�����������܂��B���������āA�����זE�́A���ȗR���̃^�� �p �N�������łȂ��A�E�C���X�R���̃^���p�N���̒f�Ђ����邱�ƂɂȂ�܂��B����ɑ��āAMHC�N���XII�́AB�זE�����זE�Ȃnj���ꂽ�זE�������� ���A�O�����荞�^���p�N���̒f�Ђ���܂��BB�זE�́AB�זE��e�̂Ɍ��������^���p�N���̒f�Ђ��悹�܂����A����זE�͐H��p�Ŏ�荞�^���p�N ���̒f�Ђ��悹�܂��B�������A����זE�ł́A��荞�^���p�N���̒f�Ђ�MHC�N���XI�ɂ��悹�邱�Ƃ��m���Ă��܂��B
�@T�זE��e�̂�[�y�v�`�h+MHC] �����̂̌����ɂ��āA�}24.6�����Ȃ���A�J��Ԃ��̕���������܂����A�������Ă����܂��傤�B
�@�w���p�[T�זE���邢�̓L���[T�זE�̕\�ʂɘI�o���Ă���T�זE��e�̂Ƃ����^���p�N���́A ���̌������ʂ�T�זE���ƂɈقȂ�܂��B���̌������ʂ̌`��1000����ވȏ゠��Ɛ��肳��܂��B�R�̂̂Ƃ��Ɠ��l�A��`�q�̍č\���ɂ�胉���_���ɂ��ꂾ ���̑��l���ݏo���Ă��܂��B���̖c��ȃ��p�[�g���[�ɂ���āA����זE������a���̗R���� [�R���y�v�`�h�{MHC] �����̂��ǂ̂悤�Ȃ��̂ł���A����Ƀs�^�b�ƌ�������T�זE��e�̂�����T�זE���قڊm���ɑ��݂��邱�ƂɂȂ�܂��i�}24.6����j�B�������A�����́AT�זE��e�� �̌������ʂ� [�R���y�v�`�h�{MHC] �����̂̌`�����ƌ����̊W�œK�����Ă��邱�ƂŋN����܂��B
�}24.6�@T�זE��e�̂� [�y�v�`�h+MHC] �����̂̌���
�@����A����זE�����ȍזE�̎��[��H�ׂāA����ɗR������ [���ȃy�v�`�h�{MHC] ����Ă��A����Ƀs�^�b�ƌ�������T�זE��e�̂�����T�זE�͂��łɔr������Ă��邽�߁A���݂���\���͂قƂ�ǂ���܂���i���̎d�g�݂ɂ��ẮA���b������ ���j�B���������āAT�זE��e�̂� [�R���y�v�`�h�{MHC] �����̂Ɍ����ł����̂́A����זE���������̂��R���y�v�`�h�ł��������Ƃ������Ă��邱�ƂɂȂ�܂��B�������A���̌����́A�P�Ɍ��ƌ����̊W�ɂ������Ƃ��������ł� ���A������������זE���p�^�[���F����e�̂ɂ��ٕ���H�ׂ��Ɣ��f���Ċ��������Ă��邱�Ƃ��O������ƂȂ��Ă���A���R�Ɖu�n�Ɗl���Ɖu�n�̃_�u���`�F�b�N �ɂ���āA���̃y�v�`�h���O���̂��̂ł���Ɣ��f���Ă��邱�Ƃɒ��ӂ��Ă����K�v������܂��B
�@�w���p�[T�זE�́A����זE������ [�R���y�v�`�h�{MHC�N���XII] �����̂Ɍ�������Ɗ���������A���B���Đ��𑝂₵�܂��B����AB�זE��B�זE��e�̂Ɍ��������^���p�N������荞�݁AMHC�N���XII���q���g���Ă��̒f�Ђ��R���� �Ă���A���������ꂽ�w���p�[T�זE��T�זE��e�̂�����Ɍ��������Ƃ���ƁAB�זE����荞�^���p�N���͈ٕ��ł��邱�Ƃ��ۏ���܂��i�}24.6�E ��j�B���Ƃ���B�זE ��e�̂́A�O���̃^���p�N���ɂ����������Ȃ��͂��ł����A�����ł��w���p�[T�זE�ɂ��_�u���`�F�b�N���A���ȗR���ł͂Ȃ����Ƃ��m�F���Ă��܂��B�������� ���n�t���ď��߂�B�זE�͊��������A���B���āA����B�זE��e�̂��R�̂Ƃ��Đ���ɎY�����A�זE�O�֕��傷��悤�ɂȂ�̂ł��B
�@�܂��A�}24.6�ɂ͂���܂��A���������ꂽ�w���p�[T�זE�͑̓������A�����ٕ���H�ׂ��}�N���t�@�[�W�Əo��ƁA���̃}�N���t�@�[�W������ ������ �Ƃ����@�\������܂��B�}�N���t�@�[�W���A[�R���y�v�`�h�{MHC�N���XII] �����̂����̍זE�\�ʂɒ��Ă��܂��B
�@����זE�́AMHC�N���X�h���g���āA�L���[T�זE�ɑ��Ă��R�����s���܂��B�L���[T�זE�́A����זE������ [�R���y�v�`�h�{MHC�N���XI] �����̂Ɍ�������Ɗ���������A���B���Đ��𑝂₵�܂��B���̊������ɂ́A���������ꂽ�w���p�[�s�זE�������Ɏ���זE�Ɍ������A���o���ꂽ�T�C�g�J�C���� �т�K�v������ꍇ������ƍl�����Ă���A��͂�_�u���`�F�b�N�̋@�\�������Ă���悤�ł��i�}24.6�����j�B����A�����זE��MHC�N���X�h���q���g�� �čR�����Ă� ��A���������ꂽ�L���[T�זE��T�זE��e�̂�����Ɍ��������Ƃ���ƁA���̍זE�������זE�ł���Ɣ��f���邱�Ƃ��ł��A���ȍזE�ł��銴���זE�ɃA�|�g�[�V �X��U�����āA�����j�܂��i�}24.6�E���j�B���̂Ƃ��A�����זE���̃E�C���X���ꏏ�ɕ�������A���ł��܂��B
�@���G�ȖƉu�V�X�e���𗝉��ł����ł��傤���B�Ɖu�זE�����Ƒ��ʂ���ɂ������āA���̑S�̑��ł͂Ȃ��A�f�Ёi�y�v�`�h�j�����ōs���Ă���Ƃ����̂͋����ł��B�R�� ���A�E�C���X�̈ꕔ�i�G�s�g�[�v�j�ւ̌����ɂ���ĔF�����Ă��܂����B�����������Ƃ��܂߁A�Ɖu�Ƃ����V�X�e���̂��_���\���͔��ɋ����[�����̂������ ���B�����Ȋw�Ƃ����͎̂����̗���ŁA������������������āA�o���邾���Ǝv���Ă������ɂ́A�����Ƃ����V�X�e���𐬂藧�����Ă���_���\���𗝉�����Ƃ��� �ʔ����������ł������Ă���������K���ł��B
�@1��ʂ��ēǂ����ł́A��������͓̂�������Ǝv���܂����A�����ł�����x�}24.3�ɖ߂� �āA�� ��܂ł̂��Ƃ����Ă݂Ă��������BT�זE��e�̂� [�y�v�`�h+MHC] �����̂̌����ɂ��ẮA����AMHC�ɒ��ڂ����b�������Ɏ��グ��\��ł��B
![]()